
di Giovanni Mian
1 Introduzione
I prodotti di IV gamma
La commercializzazione di frutta, ortaggi e verdure è oggi suddivisibile in cinque gamme, distinte dal processo di lavorazione subito dalla raccolta alla vendita e così suddivise in:
- Prima gamma: ortofrutta fresca
- Seconda gamma: ortofrutta e verdure in conserva proposte in
- Terza gamma: frutta e verdure
- Quarta gamma: ortofrutta fresca, lavata, confezionata e pronta al
- Quinta gamma: frutta e verdure cotte e ricettate, confezionate e pronte al
In particolare, con la dicitura “prodotti di IV gamma” si indicano la frutta, la verdura e, in generale, gli ortaggi freschi, a elevato contenuto di servizio, confezionati e pronti per il consumo (Fonte: www.aiipa.it).
Questi prodotti sono noti anche come prodotti minimamente trattati, pronti da mangiare (ready to eat), tagliati da poco (fresh cut), pronti da cuocere (ready to cook) (Colelli, 2009). In questa categoria, oltre alle insalate in busta e la frutta pronte per essere consumate, rientrano anche gli ortofrutticoli subito disponibili per la preparazione di alimenti da cuocere (es: verdure per minestrone).
Le tecniche di produzione fuori suolo
L’innovazione nel settore dell’ortofloricoltura ha portato allo sviluppo di pratiche che prendono il nome di “fuori suolo”, alternative alla tradizionale produzione in suolo. Questo sistema di coltivazione fu messo a punto già nel 1860, ma solo a partire dal 1929 ha cominciato ad essere utilizzato per la produzione di ortaggi su scala commerciale, grazie al fisiologo William F. Gericke.
Le tecniche di produzione fuori suolo sono suddivise in base all’utilizzo o meno del substrato. Nel primo caso si ricorre all’utilizzo di materiali che possono essere classificati a seconda della natura in: naturali organici (torba, fibra di cocco, paglia, stocchi di mais, segatura, pula di riso, alghe), naturali inorganici (lapillo, sabbia, pomice, argilla espansa, perlite, vermiculite, lana di roccia) sintetici (polistirolo, poliuretano, resina, urea formaldeide).
La produzione senza substrato, invece, viene definita più comunemente con il termine di idroponica (dal greco: ‘idros’, acqua e ‘ponos’, lavoro) e comprende tre sistemi principali: NFT (Nutrient Film Technique), aeroponia e floating system (Lazzarin et al., 2007). Nello specifico, il metodo floating system in cui le piante, sostenute da pannelli galleggianti, crescono con le radici immerse nella soluzione nutritiva per l’intero ciclo colturale Questo sistema presenta numerosi vantaggi, rispetto alla coltivazione tradizionale su terreno, sia dal punto di vista qualitativo che quantitativo: una molteplicità di applicazioni pratiche, che variano dagli studi fisiologici, alla produzione commerciale di ortaggi e fiori, alle possibilità di impiego in ambienti anomali (ad es. in Antartide, nello spazio), nell’agricoltura urbana oppure nei terreni affetti da stanchezza o da problemi fitosanitari (Tesi, 2005). Rispetto alla coltivazione in pieno campo, inoltre, la crescita è più veloce e quindi l’entrata in produzione avviene prima e si riscontra anche un aumento di produzione rispetto la coltivazione tradizionale. Inoltre, si ottengono prodotti più puliti e senza problematiche dovute a fitopatie. Per i numerosi vantaggi offerti, il floating system si è molto diffuso in Italia, dove è impiegato per la produzione di ortaggi da foglia, a cespo (es. insalata) o come “baby leaf” (es. rucola, valerianella), destinati alla IV gamma, per i quali i requisiti qualitativi sono piuttosto restrittivi.
Tra le specie più coltivate per ottenere insalata da taglio vi è la valerianella (Valerianella locusta (L.) Laterr). Dal punto di vista nutritivo è caratterizzata da un basso valore energetico (10 cal/100 g) e da un buon contenuto in vitamine e sali minerali. Negli ultimi anni il suo consumo è decisamente in crescita. Per la produzione fuori suolo della Valerianella viene generalmente adoperato il sistema del floating system, con cicli di produzione che vanno da 45 a 55 giorni.
La biofortificazione
Con il termine “biofortificazione” si intende l’aumento in elementi nutritivi (minerali o vitamine) benefici per la salute dell’uomo, nella parte edibile dei vegetali con un concomitante incremento delle caratteristiche quanti-qualitative del prodotto. I prodotti biofortificati, quindi, aiutano a prevenire l’insorgenza di problemi di salute legati alle carenze nutrizionali (Welch, 2001). Infatti la malnutrizione, ossia lo squilibrio nell’assunzione di nutrienti mediante la dieta, è un problema diffuso a livello mondiale che provoca più di 20 milioni di morti l’anno (Kennedy et al., 2003; WHO and FAO, 2003). In questo contesto, la biofortificazione è di primaria importanza, ma la sua efficacia non dipende solo dalla concentrazione di microelementi nella parte vegetale destinata al consumo umano, ma anche dalla biodisponibilità di tali nutrienti, cioè dalla quantità che il corpo umano è in grado di assumere dopo i processi di trasformazione.
Il Selenio e la sua importanza per la salute umana
Il Selenio (Se) è un micronutriente molto studiato sia in ambito medico, per le notevoli implicazioni che ha nella salute umana, che in ambito vegetale per le ripercussioni che un suo accumulo può avere nella fisiologia della pianta e nella nutrizione di animali ed esseri umani.
Nell’organismo umano il Se è un importante micronutriente. È considerato, infatti, un ottimo antiossidante in grado di cooperare anche con altri elementi quali le vitamine C ed E, ed esplicare un ruolo di protezione dai radicali liberi (Mach, 2008). Del Se è noto anche il coinvolgimento nel metabolismo degli ormoni tiroidei, nel sistema immunitario, nell’inibizione e contrasto delle malattie virulente, nel ridurre lo sviluppo dell’AIDS attraverso una diminuzione della velocità di sviluppo del virus HIV e nella soppressione del carcinoma (Rayman, 2000). Non solo, tale elemento, attraverso un apporto equilibrato mediante la dieta, aiuta alcuni casi di complicazione in soggetti afflitti da diabete. Funge da elemento di prevenzione nei confronti dell’asma e, attraverso l’inibizione dei radicali liberi, modula l’effetto delle radiazioni. Inoltre, svolge un ruolo fondamentale connesso al funzionamento dei neurotrasmettitori cerebrali (le sue carenze velocizzano la demenza senile dovuta ad Alzheimer). È stato osservato anche che, in alcune aree della Cina in cui le diete sono scarse di Se, è maggiore l’insorgenza di malattie come la cardiomiopatia e l’osteoporosi (Surai et al., 2003).
Presente in due diversi aminoacidi, la Se-cisteina (Se-Cys) e la Se-metionina (Se-Met), è un importante componente delle cosiddette selenoproteine, che esplicano funzioni fondamentali nell’organismo umano. La Se-Cys è incorporata all’interno delle proteine attraverso uno specifico meccanismo. Questo particolare aminoacido è codificato dal codone UGA nell’mRNA, che è letto normalmente come codone di terminazione della traduzione, ma in specifici contesti determina l’inserimento delle Se-Cys nella catena aminoacidica delle selenoproteine (Boeck et al., 1991). La più importante e conosciuta Se-proteina nel corpo umano è la glutatione perossidasi, implicata nella difesa dagli stress ossidativi indotti dai radicali liberi. Altre sono la tioredossina reduttasi, implicata nel mantenimento dello stato redox intracellulare, la iodotironina deiodinasi implicata nel metabolismo degli ormoni tiroidei. Ulteriore esempio di Se-proteine sono quelle che mantengono l’integrità del flagello nello spermatozoo (Brown e Arthur, 2001).
La principale fonte di Se per l’uomo si trova nel cibo di origine vegetale, in cui l’elemento si può trovare in quantitativi e forme chimiche (sia inorganiche, sia organiche) diversi (Pedrero e Madrid, 2009). Il metabolismo del Se nell’organismo umano dipende principalmente dalla forma in cui sitrova: ad esempio, la Se-Cys e la Se-Met sono forme prontamente disponibili in quanto facilmente incorporate nella catena aminoacidica delle selenoproteine. Atre forme chimiche possono avere effetti benefici sulla salute. È stato osservato che i componenti metilati sono importanti per la prevenzione dal cancro e non sono coinvolti nella proteinogenesi, come la seleno- metil- selenocisteina e i derivati del γ-glutamile (trovati in piante di Allium e Brassica) (Böck et al., 1991).
Il selenio nelle piante: importanza, assorbimento e metabolismo
L’essenzialità del selenio nelle piante è tuttora in discussione, sebbene ci siano evidenze di una sua funzione benefica. Infatti, a basse concentrazioni, funge da antiossidante, esercita un ruolo di protezione nei confronti delle cellule incrementando la resistenza ai raggi UV, ritarda la senescenza incrementando i livelli della glutatione perossidasi, promuove la crescita delle plantule; l’assorbimento e l’assimilazione dell’azoto ed inoltre è in grado di regolare lo stato idrico della pianta (Hartikainen e Xue, 1999; Xue et al., 2001). L’accumulo del Se nei vegetali dipende principalmente dalla forma chimica in cui si trova e da fattori come il pH, la presenza di carbonato di Ca e dalla concentrazione dello stesso. Il Se entra come selenato (principalmente) o selenito nelle cellule delle radici attraverso un processo di trasporto attivo, mediato dai trasportatori dello ione solfato. In particolare, sono coinvolte proteine denominate SULTR (sulphate transporters) che presentano alta affinità per lo zolfo e mediano il trasporto in simporto con tre protoni.
Lo S e il Se, essendo trasportati dalle stesse proteine di membrana, si trovano in competizione. Infatti, l’eccessiva disponibilità di solfato inibisce l’accumulo di Se. Non tutte le specie vegetali rispondono allo stesso modo. Le piante Se-accumulatrici, ad esempio, assorbono preferenzialmente il Se piuttosto che lo S (Wu e Huang, 1991). La traslocazione del Se dalle radici alle foglie dipende principalmente dalla forma in cui l’elemento viene assorbito. Il selenato, per esempio, è trasportato molto più facilmente del selenito e delle forme organiche come la Se- metionina (Clarkson e Luttge, 1991; Leggett e Epstein, 1956).
Successivamente al trasporto all’interno delle cellule radicali, il selenato è traslocato attraverso lo xilema verso le foglie. Il selenato entra nei cloroplasti, dove è metabolizzato mediante la via di assimilazione riduttiva del solfato. Il primo passo consiste nella sua attivazione ad opera dell’enzima ATP sulfurilasi (ATPS) con formazione di adenosinfosfoselenato (APSe). Seguono diversi step di riduzione, che coinvolgono l’APS riduttasi (APR) e la sulfito riduttasi (SiR) con formazione di seleniuro (Se2-). L’APS può essere ridotto anche attraverso una fase non enzimatica ad opera della GSH e una enzimatica catalizzata dalla GSH reduttasi con formazione di seleniuro coniugato a glutatione (GS-Se-). L’ultima fase della via di assimilazione consiste nell’incorporazione del seleniuro nell’OAS, reazione catalizzata dalla O-acetilserina tioliasi (OASTL). L’OAS è prodotta in seguito al trasferimento di un gruppo acetilico donato dall’acetil- coenzima A nella serina per opera dell’acetilserina transferasi (SAT). SAT e OASTL costituiscono un unico complesso enzimatico denominato cisteina sintasi (CS) (Terry et al., 2000) (Fig. 1).
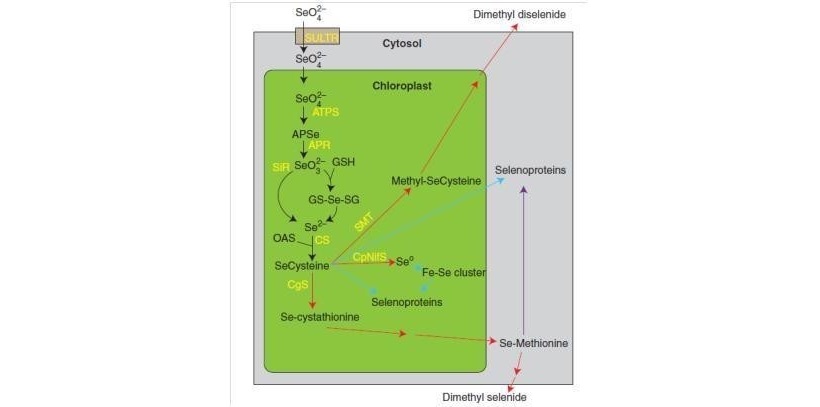
Figura 1. – Via di assimilazione del Se all’interno dei cloroplasti.
2 Scopo del lavoro
La qualità è un aspetto sempre più richiesto dal consumatore, soprattutto nel settore della IV gamma, che negli ultimi anni ha subito un forte incremento, interessando una fetta di mercato crescente.
Scopo della tesi è quindi stato di valutare la possibilità di biofortificare la Valerianella locusta (L.) allevando piante di due diverse cultivar (Baron e Gala), con un sistema idroponico che simuli il floating system, in presenza di selenio a diverse concentrazioni. A questo scopo sono stati analizzati i parametri di crescita quali l’accumulo di peso fresco in foglie e radici e il contenuto di clorofilla (indice SPAD). Inoltre, data l’interazione tra il Se e lo S, è stata valutata l’attività degli enzimi ATP sulfurilasi e O-acetilserina-tioliasi rispettivamente primo e ultimo enzima nella via di assimilazione dello zolfo e del selenio.
3 Materiali e metodi
Le analisi sono state condotte su due diverse cultivar di valerianella (Valerianella locusta (L.) Laterr),‘Baron’ e ‘Gala’, coltivate mediante sistema di coltivazione fuori suolo, ossia il floating system (Fig. 2). Sono stati valutati tre diversi trattamenti: 0 (controllo), 10 e 40 μM di Se (il selenio è stato aggiunto sotto forma di Na2SeO4). La soluzione nutritiva è stata rinnovata con cadenza settimanale. L’intero ciclo di coltivazione è durato complessivamente 45 giorni.

Fig. 2 – Vasche di coltivazione
Al termine del ciclo di coltivazione è stato misurato il valore di SPAD, indice del contenuto relativo di clorofilla, utilizzando un misuratore portatile “SPAD-502” (Minolta Corp, Ramsey, N.Y. USA). Inoltre, è stata misurata la resa in termini di produzione di biomassa fresca. Sono state raccolte le foglie e le radici ed è stato misurato il peso fresco mediante l’utilizzo di una bilancia analitica. Dal materiale raccolto è stato isolato un estratto proteico con lo scopo di misurare l’attività enzimatica di due enzimi coinvolti nella via di assimilazione dello zolfo, l’ATP sulfurilasi e l’O– acetilserina (tiol) liasi. Alla fine è stata condotta l’analisi statistica dei dati con il modello dell’ANOVA con cui è stata verificata l’omogeneità della varianza, secondo un valore di probabilità (P) pari a 0,05. Q
Nei grafici sono riportate le medie ± la deviazione standard di 4 repliche biologiche (N = 4); le medie che presentano differenze significative sono inoltre indicate da lettere diverse.
4 RISULTATI
Effetto del Selenio sulla produttività
Al termine del ciclo di coltivazione, per ciascuna replica sono stati registrati i pesi relativi all’apparato fogliare (parte edule) e radicale al fine di verificare l’effetto esercitato dalle diverse concentrazioni di selenio sulla produttività della valerianella.
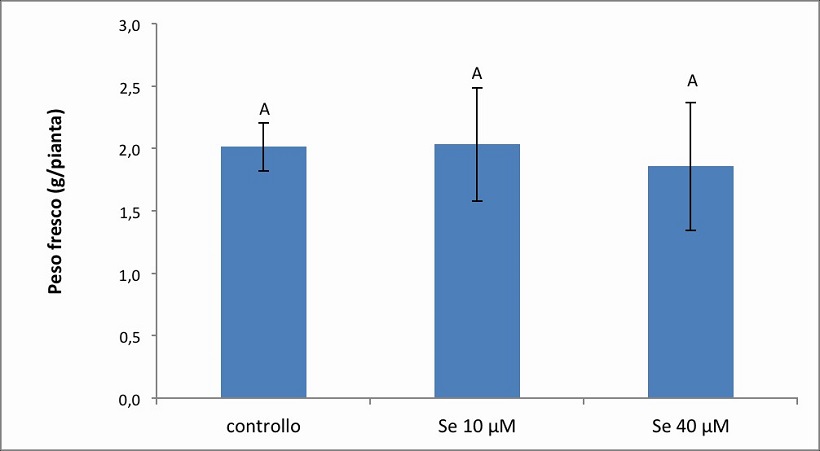
Figura 3. – Produttività espressa in g/pianta dell’apparato fogliare della cv. Gala. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nella cv. Gala (Figura 3) la misura della biomassa fogliare è risultata pari a 2.0139 g, 2.0343 g e 1.8582 g nel controllo, nel trattamento 10 µM e 40 µM di Se, rispettivamente. Non sono state osservate differenze significative nelle tre diverse condizioni di crescita, se non una leggera diminuzione, pari al 8 %, alla concentrazione 40 µM di Se, rispetto al controllo.
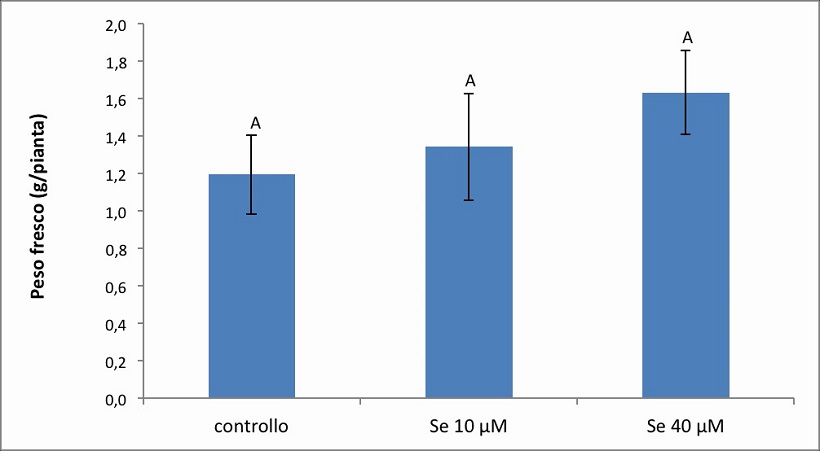
Figura 4. – Produttività espressa in g/pianta dell’apparato fogliare della cv. Baron. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Anche nella cv. Baron la misura del peso fresco fogliare (Figura 4) non ha mostrato sostanziali differenze, in termini di biomassa prodotta, tra le piante coltivate in presenza di Se e il controllo. Tuttavia è stata osservata una tendenza all’incremento del peso fresco a seguito del trattamento con Se: il peso fresco nel controllo, e nei due trattamenti 10 e 40 µM di Se, è risultato infatti pari rispettivamente a 1.1946 g, 1.3428 g e 1.6325 g. Dal confronto con il controllo quindi si evidenzia un aumento di produttività del 12% (10 µM di Se) e del 36% (40 µM di Se), rispettivamente.
Dall’analisi statistica è emerso una significatività dell’effetto semplice “cv”, mentre per l’effetto semplice Se e dell’interazione “Se per cv” non si riscontra significatività.
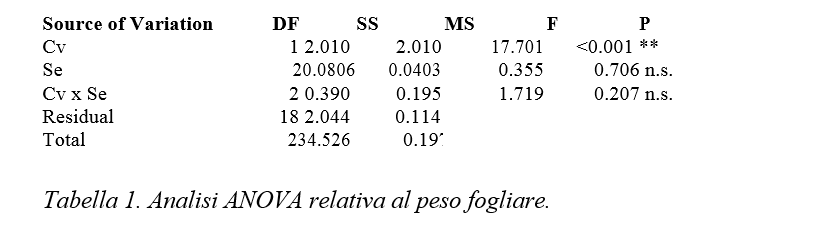
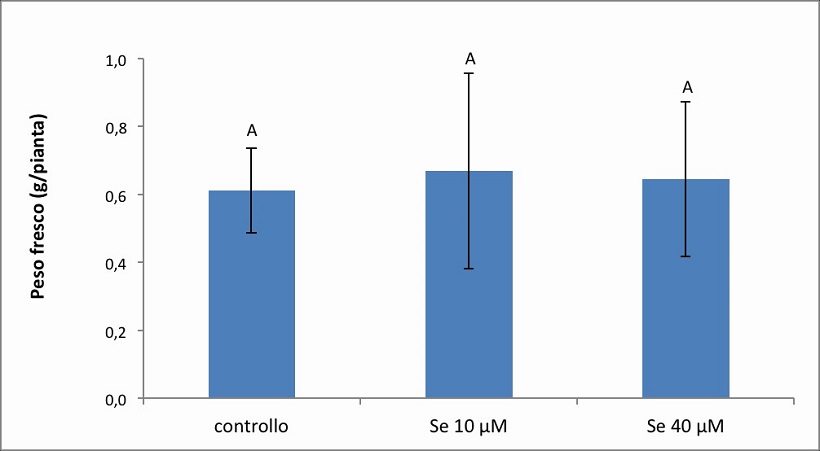
Figura 5. – Produttività espressa in g/pianta dell’apparato radicale della cv. Gala. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nella cv. Gala (Figura 5) il peso radicale è risultato mediamente pari a 0.6126 g, 0.6695 g e 0.6454 g nel controllo, Se 10 µM e Se 40 µM, rispettivamente. In modo analogo a quanto osservato per l’apparato fogliare, il peso medio delle radici non ha mostrato differenze significative nei tre diversi trattamenti. Infatti, rispetto al controllo, è stato osservato solamente un aumento del 9 % a 10 µM di Se e del 4 % a 40 µM di Se.
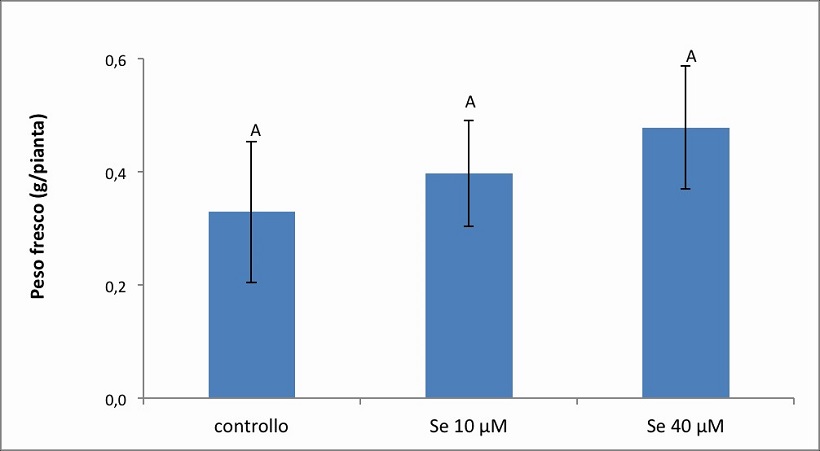
Figura 6. – Produttività espressa in g/pianta dell’apparato radicale della cv. Baron. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nella cv. Baron (Figura 6) la biomassa radicale è risultata mediamente pari a 0.3293 g nel controllo, 0.3976 g nel trattamento 10 µM di Se e 0.4780 nel trattamento 40 µM di Se. Come per la parte aerea, l’aggiunta di selenio alla soluzione nutritiva ha determinato un tendenziale incremento del peso fresco radicale rispetto al controllo. In particolare, è stato osservato un aumento di circa il 20 % nel trattamento con 10 µM di Se e un incremento più rilevante del 45 % nel trattamento con 40 µM di Se.
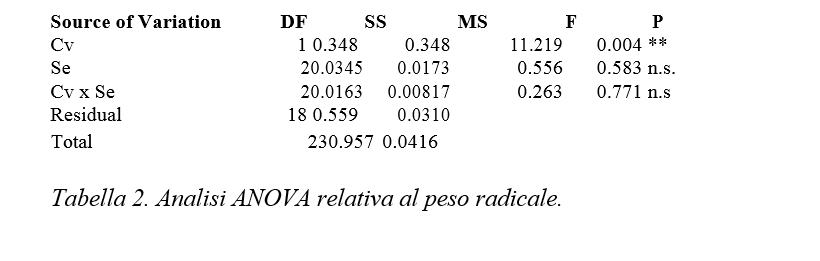
Dall’analisi statistica risulta significativo l’effetto semplice “cv”, mentre non sono significativi nè l’effetto semplice Se nè l’interazione “cv per Se” (Tabella 2).
Effetto del Selenio sull’indice SPAD
L’analisi del contenuto relativo di clorofilla, valutato misurando il valore dell’indice SPAD, ha evidenziato ulteriori differenze tra i due genotipi considerati.
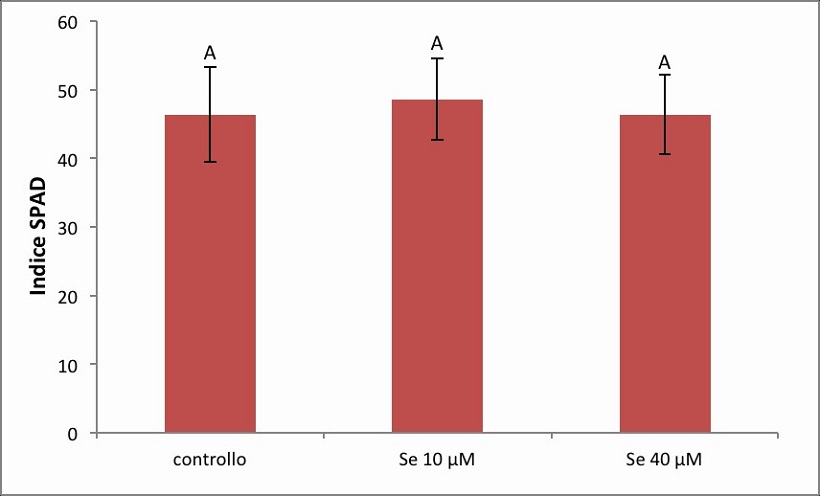
Figura 7. – Indice SPAD relativo alla cv Gala. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Dall’analisi dell’indice SPAD misurato nella cv. Gala (Figura 7) non sono emerse differenze significative tra le tre condizioni di crescita, con valori medi nel controllo, trattato 10 µM di Se e trattato 40 µM di Se pari rispettivamente a 46.29, 48.56 e 46.33 unità.
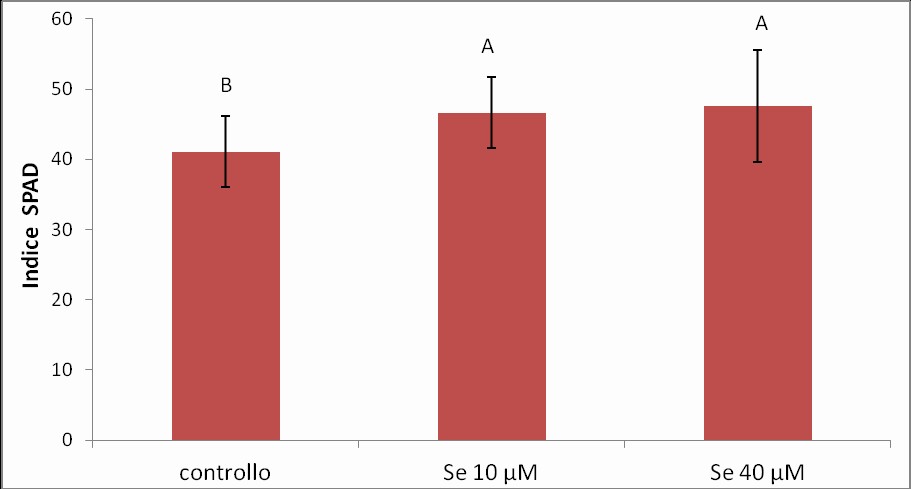
Figura 8. – Indice SPAD relativo alla cv. Baron. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nella cv. Baron (Figura 8) è stato invece osservato un incremento progressivo del contenuto relativo di clorofilla all’aumentare della disponibilità di Se. Il valore medio dell’indice SPAD è risultato significativamente più elevato nelle piante trattate con Se 10 µM (46.63) e 40 µM (47.57) rispetto al controllo (41.11).
Dall’analisi statistica emerge significatività sia per gli effetti semplici “cv” e “Se” sia per l’interazione “cv per Se” (Tabella 3).

Effetto del Selenio sull’attività dell’ATP sulfurilasi (ATPS)
I valori dell’attività dell’enzima ATP sulfurilasi rilevati nelle foglie delle due cv. di valerianella sono riportati nella Figura 9, mentre i dati relativi alle radici sono presentati nella Figura 10.
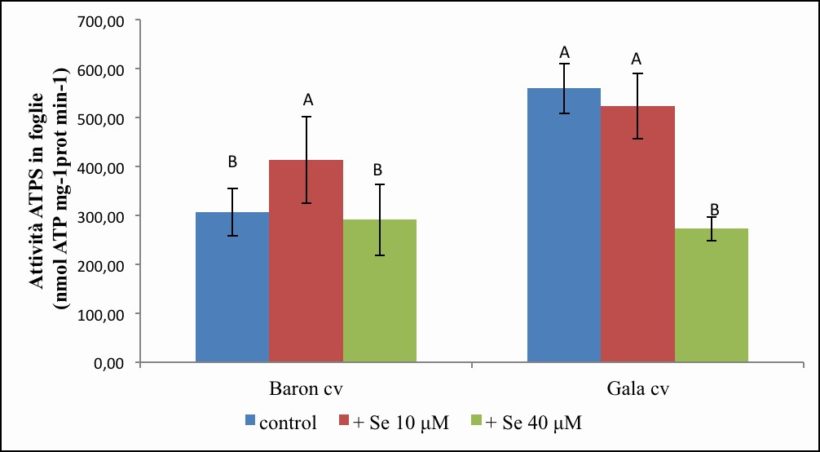
Figura 9. – Attività dell’ATPS in foglie espressa in nmol ATP prodotto x mg-1 proteina x min-1. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
L’analisi di questo enzima nelle foglie della cv. Baron (Figura 9) ha evidenziato delle attività mediamente pari a 306.67, 412.68 e 290.37 nmol ATP x mg-1 proteina x min-1, rispettivamente nel controllo e nei trattamenti a 10 e 40 µM di Se. In particolare, si osserva un incremento significativo nelle foglie trattate con 10 µM di Se rispetto al controllo (+ 34 %), mentre nelle piante trattate con 40 µM di Se non si evidenziano sostanziali variazioni.
Per quanto riguarda la cv. Gala (Figura 9) si osserva un’attività nelle piante di controllo maggiore di quella misurata nella cv. Baron, con valori mediamente pari a 559.12 nmol ATP x mg-1 proteina x min-1. IL trattamento con 10 µM di Se non comporta variazioni significative (523.23 nmol ATP x mg- 1 proteina x min-1), mentre a 40 µM di Se si osserva una diminuzione consistente (- 52 %) e significativa dell’attività enzimatica rispetto al controllo, che raggiunge valori medi pari a 272.25 nmol ATP x mg-1 proteina x min-1.
Dall’analisi statistica si evidenzia una significatività sia per gli effetti semplici “Se” e “cv” che per l’interazione “Se per cv” (Tabella 4).

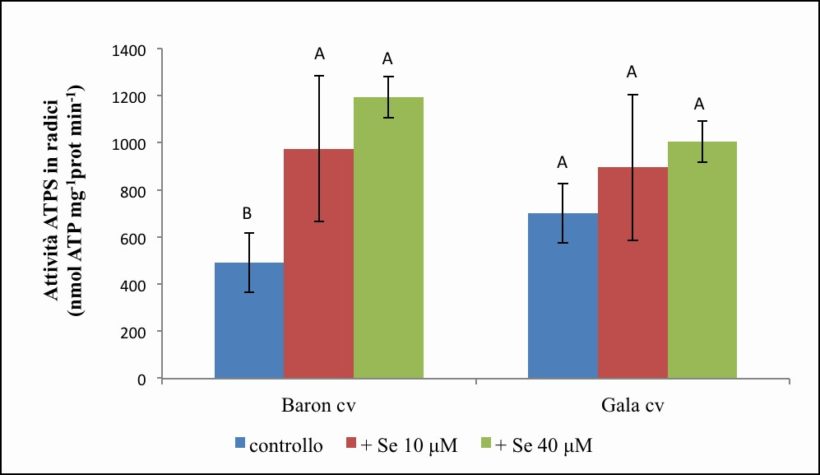
Figura 10. – Attività dell’ATPS in radici misurata in nmol ATP x mg-1 proteine x min-1. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
L’attività dell’enzima risulta maggiore nelle radici rispetto alle foglie, indipendentemente dal trattamento del selenio. Rispetto al controllo (491.54 nmol ATP x mg-1 proteina x min-1), la somministrazione di Se determina nelle radici della cv. Baron (Figura 10) un significativo incremento di attività, con valori medi di 973.82 (+ 98 %) e 1194.21 (+ 142 %) nmol ATP x mg-1 proteina x min- 1, rispettivamente a 10 µM e 40 µM di Se.
Per quanto concerne la cv. Gala (Figura 10), l’attività misurata nel controllo è mediamente pari a 701.74 nmol ATP x mg-1 proteina x min-1, e aumenta, seppur non significativamente, nelle radici trattate con 10 µM di Se (895.42 nmol ATP x mg-1 proteina x min-1, + 27 %) e 40 µM di Se (1005.78 nmol ATP x mg-1 proteina x min-1, + 43 %).
Dall’analisi statistica si evince un effetto significativo solo per l’effetto semplice “Se”, mentre per l’effetto semplice “cv” e per l’interazione “cv per Se” non emerge significatività (Tabella 5).

Effetto del Selenio sull’attività O-acetilserina-tioliasi (OASTL)
I valori dell’attività dell’enzima O-acetilserina-tioliasi rilevati nelle foglie delle due cv. di valerianella sono riportati nella Figura 11, mentre i dati relativi alle radici sono presentati nella Figura 12.
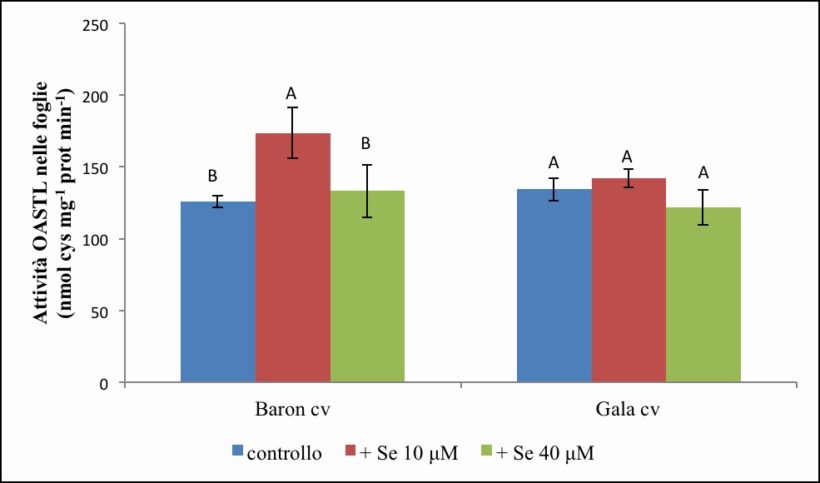
Figura 11. – Attività dell’OASTL in foglie misurata in nmol cys x mg-1 prot x min-1. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nella cv. Baron (Figura 11) l’attività dell’OASTL fogliare è risultata pari mediamente a 125.91 nmol cys x mg-1 prot x min-1 nel controllo; raggiunge valori significativamente più elevati del 37 % (173.56 nmol cys x mg-1 prot x min-1) nelle piante trattate con 10 µM di Se, mentre torna a valori simili al controllo nelle piante trattate con 40 µM (133.25 nmol cys x mg-1 prot x min-1).
Per la cv. Gala (Figura 11), l’attività enzimatica mostra valori medi di 134.47 nmol cys x mg-1 prot x min-1 nel controllo e non varia significativamente a seguito del trattamento con Se (142.04 e 121.99 nmol cys x mg-1 prot x min-1 nei trattamenti a 10 µM e 40 µM di Se, rispettivamente).
Dall’analisi statistica si evidenzia una significatività per gli effetti semplici “cv”, “Se” e per l’interazione “cv per Se” (tabella 6).
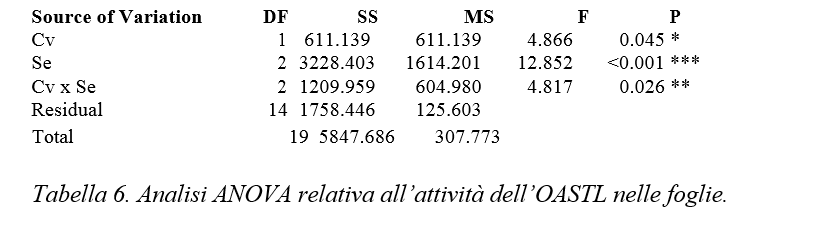
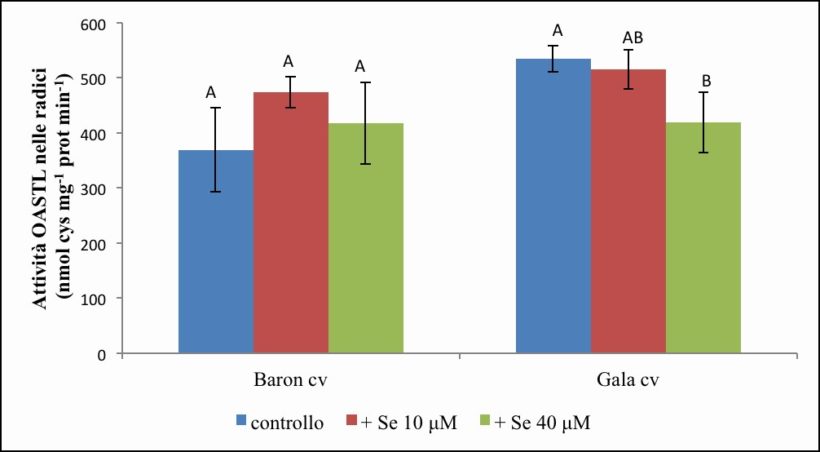
Figura 12. – Attività dell’OASTL in radici misurata in nmol cys x mg-1 prot x min-1. I valori indicano medie ± deviazione standard di 4 repliche biologiche. Lettere diverse indicano differenze significative tra le medie dei trattamenti (ANOVA, P<0,05, N=4).
Nelle radici della cv. Baron (Figura 12) l’attività enzimatica è risultata pari mediamente a 369.54 nmol cys x mg-1 prot x min-1 nel controllo, 473.43 nmol cys x mg-1 prot x min-1 nel trattamento con Se 10 µM e 417.4 nmol cys x mg-1 prot x min-1 in quello con Se 40 µM. Rispetto al controllo si evidenzia un tendenziale aumento di attività del 28 % alla concentrazione 10 µM di Se e un successivo calo alla concentrazione 40 µM di Se.
Per quanto concerne la cv. Gala (Figura 12), l’attività di base nelle radici non trattate è leggermente maggiore rispetto a quella misurata per la cv. Baron ed è pari mediamente a 534.92 nmol cys x mg-1 prot x min-1 nel controllo e a 515.54 e 418.91 nmol cys x mg-1 prot x min-1 nei campioni trattati rispettivamente con Se 10 µM e 40 µM. Questi risultati mettono in evidenza una tendenziale diminuzione dell’attività enzimatica per effetto della somministrazione di Se.
Dall’analisi statistica emerge significatività per entrambi gli effetti semplici “cv” e “Se” e per la loro interazione (Tabella 7)
5 DISCUSSIONE
I prodotti di IV gamma sono una componente alimentare che agevola il nostro stile di vita. Permettono, infatti, di aver sempre a disposizione un prodotto fresco e prontamente utilizzabile che riduce la tempistica di preparazione dei pasti, aspetto di notevole rilevanza nella caotica società odierna. Dato l’aumentato consumo di questi alimenti e il ruolo che svolge la dieta sulla salute dell’uomo, assumono fondamentale importanza le caratteristiche di qualità. In questo contesto, il presente lavoro si è focalizzato sul miglioramento, quantitativo e qualitativo, della coltivazione fuori- suolo di valerianella destinata ai prodotti di IV gamma intervenendo sulla composizione della soluzione nutritiva. In particolare, l’attenzione è stata rivolta alla biofortificazione con selenio, elemento essenziale per la salute dell’uomo in quanto componente fondamentale di alcuni enzimi chiave, quali glutatione perossidasi (funzione antiossidante), tioredossina reduttasi (mantenimento dello stato redox intracellulare), iodotironina deiodinasi (metabolismo ormoni tiroidei), e di proteine quali le Se-proteine implicate nel mantenimento dell’integrità del flagello nello spermatozoo (Brown e Arthur, 2001). Gli esperimenti condotti sono stati finalizzati alla valutazione della resa di produzione e dello stato nutrizionale della pianta mediante stima del contenuto di clorofilla e misura dell’attività di due enzimi chiave coinvolti nel metabolismo dello zolfo (indicativo dell’attività di assimilazione del selenio in forme organiche); per questo il confronto è stato eseguito tra piante di due cultivar differenti allevate in condizioni standard e piante coltivate in presenza di Se a due diverse concentrazioni, 10 e 40 µM.
In generale, le analisi effettuate hanno evidenziato una diversa risposta delle due cultivar alla presenza di Se. Queste differenze possono essere attribuite a una diversa capacità di accumulare e metabolizzare questo elemento.
Il primo aspetto esaminato è stato il contenuto relativo di clorofilla come indice indiretto dello stato nutrizionale della pianta. Nella cv. Baron è stato riscontrato un incremento progressivo della quantità di pigmenti fotosintetici all’aumentare della disponibilità di Se, contrariamente alla cv. Gala in cui non sono state osservate variazioni significative. L’effetto positivo sulla crescita delle piante trova conferma nella misura della biomassa prodotta, valutata al termine del ciclo di coltivazione. In primo luogo è stata considerata la parte edule del prodotto, fattore determinante nell’ottica del produttore. Solamente la cv. Baron ha risposto positivamente alla presenza di Se. È stato osservato, infatti, un progressivo incremento del peso fresco all’aumentare della disponibilità del Se. Nella cv. Gala, invece, non sono stati osservati sostanziali cambiamenti nelle piante trattate rispetto al controllo. Il Se quindi, somministrato a determinate concentrazioni, non solo non risulta tossico ma può anche esercitare un effetto positivo sulla crescita della valerianella. Simili risultati sono stati osservati in Lactuca Sativa coltivata con sistema fuori suolo in presenza di concentrazioni inferiori a 15 µM di Se (Hawrylak-Nowak, 2013). Un aumento di biomassa e una miglior colorazione della parte edule rappresentano aspetti di particolare interesse per il mercato di questi prodotti.
Oltre a valutare la risposta della pianta in termini di produzione è stato considerato anche l’effetto della somministrazione del selenio sul metabolismo. In particolare sono stati considerati due enzimi chiave, l’ATP sulfurilasi e l’O-acetilserina tioliasi, che catalizzano, rispettivamente, il primo e l’ultimo step nelle vie di assimilazione dello zolfo e del selenio. E’ noto, infatti, che i due elementi hanno proprietà chimiche molto simili e interagiscono condividendo le stesse vie di assorbimento e assimilazione.
In primo luogo è stata valutata l’attività dell’ATP sulfurilasi (ATPS), enzima che catalizza l’attivazione del substrato, sia esso solfato o selenato, step fondamentale affinché l’elemento possa poi essere assimilato. Analizzando l’attività a livello fogliare è stato osservato che nella cv. Baron il Se ha significativamente stimolato l’attività dell’ATPS solo quando fornito alla concentrazione 10 µM, nella cv. Gala invece l’attività enzimatica del controllo era superiore rispetto alla cv. Baron ma è risultata fortemente inibita dalla somministrazione di 40 µM Se. Questo comportamento suggerisce che alle concentrazioni di Se più elevate l’attività dell’enzima diminuisca per evitare un’eccessiva incorporazione di Se negli amminoacidi e ciò sarebbe coerente con l’accumulo di solfato, traslocato dalle radici, che si ritrova nelle foglie di piante trattate con 40 µM Se (Tomasi et al., 2015).
A livello radicale è stato invece osservato, per entrambe le cultivar, un aumento progressivo dell’attività dell’ATPS a concentrazioni crescenti di Se, con livelli superiori nella cv. Baron. Inoltre è emersa un’attività maggiore nel tessuto radicale rispetto a quello fogliare. Questi risultati suggeriscono un ruolo importante delle radici nell’assimilazione del solfato sia quando questo è fornito a elevate concentrazioni, come quelle comunemente utilizzate nella coltivazione idroponica, sia quando le piante sono allevate in presenza di selenio.
L’O-acetilserina-tioliasi (OASTL) è un altro enzima che svolge un ruolo importante nell’assimilazione dello S e del Se in molecole organiche. In V. locusta sono state osservate anche in questo caso differenze nei due genotipi considerati. Nella cv. Baron, le variazioni di attività hanno riproposto l’andamento osservato per l’ATPS con un incremento, sia pur non significativo a 10 µM di Se, mentre nessuna differenza è stata osservata per la cv. Gala. Questo comportamento suggerisce una minor sensibilità di questo enzima al trattamento con il Se che garantirebbe comunque il mantenimento di concentrazioni adeguate di cisteina e metionina, come dimostrato da Tomasi et al. (2015).
Per questo enzima non sono state osservate differenze nella risposta al selenio tra le foglie e le radici. Tuttavia, diversamente da quanto atteso, anche l’attività dell’OASTL è risultata maggiore nelle radici che nelle foglie. Anche in questo caso, l’alta concentrazione di zolfo nella soluzione nutritiva potrebbe favorire la via di assimilazione dello zolfo a livello radicale. L’attività di questo enzima risente comunque della disponibilità di O-acetilserina (OAS), che collega il metabolismo dello zolfo con quello di azoto e carbonio (Davidian et al., 2000).
In conclusione i risultati, ottenuti dal presente lavoro condotto su V. locusta allevata con sistema di coltivazione floating system, hanno evidenziato l’efficacia della somministrazione di Se alla soluzione nutritiva, dimostrando un miglioramento dei parametri quantitativi e qualitativi. Tuttavia gli effetti esercitati dal Se devono essere sempre considerati in relazione al genotipo e alla quantità somministrata. Se utilizzato alla giusta concentrazione il Se può anche promuovere l’assorbimento e l’assimilazione dello zolfo. Ricerche condotte sull’interazione tra Se e S in Arabidopsis thaliana e suoi mutanti hanno dimostrato che la maggiore disponibilità di Se promuove l’accumulo di S nelle foglie, dato correlato anche alla maggior attività dei trasportatori SULTR (White et al., 2004). Queste evidenze sono state confermate da studi più recenti (Ramos et al., 2010). In valerianella è stato dimostrato che il trattamento con 10 µM di Se determina l’aumento di zolfo, cisteina e metionina, oltre che la sintesi di Se-amminoacidi (Tomasi et al., 2015).
La possibilità di produrre colture biofortificate con Se può portare a un maggior valore nutrizionale del prodotto e a una sua valorizzazione commerciale. Dato che il Se ritarda la senescenza delle piante, prolungandone quindi la durata, attraverso la biofortificazione potrebbe essere anche incrementata la shelf life dei prodotti di IV gamma.
Giovanni Mian si è diplomato all’Istituto tecnico agrario “P. d’Aquileia” di Cividale del Friuli Ha conseguito la laurea triennale in Scienze agrarie presso l’Università degli studi di Udine. Successivamente si è iscritto, nello stesso ateneo, al corso di laurea magistrale in Scienze e tecnologie agrarie, laureandosi ad ottobre del 2017 con il massimo dei voti. Vanta numerose esperienze lavorative nel settore agricolo e nel settore della sperimentazione di molecole usate come fitofarmaci. E-mail: giovanni.mian@outlook.it





")