
di Giuseppe Accomando
Gregorio Mendel e le sue scoperte
Gregorio Mendel, biologo austriaco (Slesia 1822 – Brno Moravia 1884) frate agostiniano del convento di Brno, insegnante di Fisica e Scienze naturale, fu il primo che studiò il comportamento ereditario dei caratteri qualitativi, detti anche mendeliani, non facilmente misurabili a variabilità discontinua, influenzati da pochi geni o da una coppia di geni allelomorfi, caratteri definiti attraverso un attributo: colore, presenza o assenza delle corna, forma, etc.
Mendel tra il 1857 e il 1868 si dedicò ad esperienze di ibridazione tra piante del genere Pisum nell’orto del convento, e pubblicò le sue esperienze nel 1866 – 1869 “Saggio sugli ibridi vegetali”. Gli studi rimasero ignorati fino al 1900 quando, ad opera di tre scienziati (Correns, Tschermak e De Vries), rispettivamente di nazionalità tedesca, austriaca e olandese, furono divulgati al mondo scientifico come leggi di Mendel.
 Gregorio Mendel (fonte Wikimedia.org)
Gregorio Mendel (fonte Wikimedia.org)
Il merito di Mendel non fu solo quello di osservare il comportamento dei caratteri nelle generazioni, ma anche il tenere conto dei rapporti statistici con i quali i caratteri si manifestano nella massa delle piante allevate. Prima di iniziare gli incroci egli ottenne, tramite l’autofecondazione delle piante, le linee pure, che chiamò generazione parentale (P) piante omozigote rispetto ad un carattere, e cominciò dapprima prendendo in considerazione un solo carattere, poi due e così di seguito, enunciando le sue esperienze nelle famose tre leggi.
1a Legge – Dominanza o della uniformità degli ibridi in F1
2a Legge – Segregazione o disgiunzione dei caratteri in F2
3a Legge – Indipendenza dei caratteri
1a Legge: dominanza dei caratteri o uniformità degli ibridi F1
Tutti i figli derivati dall’unione fra due individui, che differiscono per un solo carattere, sono uguali tra loro; esempio se si incrociano due bovini di cui uno presenta il mantello nero uniforme e l’altro mantello rosso uniforme, in F1 (1a generazione filiale) tutti gli individui presentano il mantello nero uniforme, perché il colore nero è carattere dominante rispetto al rosso che è recessivo.
 In questi casi l’individuo riceve dai genitori per lo stesso carattere lo stesso gene, un gene è trasmesso dal cromosoma paterno, l’altro gene è trasmesso dal cromosoma materno.
In questi casi l’individuo riceve dai genitori per lo stesso carattere lo stesso gene, un gene è trasmesso dal cromosoma paterno, l’altro gene è trasmesso dal cromosoma materno.
2a Legge: segregazione o disgiunzione dei caratteri F2
I figli derivati dall’incrocio degli individui della 1a generazione F1 non sono tutti geneticamente uguali, infatti, incrociando il prodotto della F1 si hanno, nella seconda generazione filiale F2, quattro possibili combinazioni, perché c’è la separazione degli alleli; perciò, si ha, la probabilità che il 25 % siano omozigoti dominanti, il 25 % omozigoti recessivi, il 50 % eterozigoti.
 Mendel per rilevare lo stato di omozigosi o di eterozigosi degli individui che manifestavano lo stesso fenotipo incrociò a caso una pianta con fenotipo dominante con una che manifestava il carattere recessivo, nella discendenza se gli individui presentavano per il 50% il carattere dominante e per il restante 50% il carattere recessivo concludeva che la pianta madre presa a caso era eterozigote Aa , viceversa se la pianta madre era omozigote dominante AA incrcociandola con la pianta recessiva omozigote aa tutti i discendenti manifestavano solo il carattere dominante Aa fenotipo nero uniforme.
Mendel per rilevare lo stato di omozigosi o di eterozigosi degli individui che manifestavano lo stesso fenotipo incrociò a caso una pianta con fenotipo dominante con una che manifestava il carattere recessivo, nella discendenza se gli individui presentavano per il 50% il carattere dominante e per il restante 50% il carattere recessivo concludeva che la pianta madre presa a caso era eterozigote Aa , viceversa se la pianta madre era omozigote dominante AA incrcociandola con la pianta recessiva omozigote aa tutti i discendenti manifestavano solo il carattere dominante Aa fenotipo nero uniforme.
3a Legge – indipendenza dei caratteri
I geni vengono trasmessi indipendentemente gli uni dagli altri quando i caratteri presi in esame nei genitori sono più di uno, per cui si hanno nuovi fenotipi e nuovi genotipi, secondo una distribuzione statistica determinata dallo svolgimento del binomio ( 3 + 1 ) n, dove (n) indica il numero dei caratteri per cui i due individui differiscono.
Nel caso del Diibridismo (gli individui che differiscono per due caratteri ), si avrà:
 I fenotipi sono 4 così distinti:
I fenotipi sono 4 così distinti:
– 9) Nero uniforme
– 1) Rosso pezzato
– 3) Nero pezzato
– 3) Rosso uniforme
Nel Triibridismo, individui che differiscono per tre caratteri, si avrà:

Eredità dei caratteri quantitativi – Resa al Macello
I caratteri quantitativi quelli a variabilità continua, facilmente misurabili, sono di grande interesse zootecnico dal momento che caratterizzano gran parte delle produzioni economiche.
Un carattere quantitativo è determinato da una serie di loci perciò è detto anche polifattoriale o poligenico; l’espressione fenotipica, cioè la manifestazione di questi caratteri, è soggetta a influenze ambientali, particolarmente dall’alimentazione, tra questi caratteri la pelle è influenzata solo in parte dalle condizioni ambientali, il peso e lo spessore presentano una certa importanza rispetto alle attitudini produttive, nei bovini una pelle sottile è correlata positivamente con la produzione del latte e in tutti gli animali da carne influenza direttamente la resa al macello intesa come rapporto percentuale tra il peso morto (PM) dell’animale (4/4) e il peso vivo (PV) dello stesso. Il Peso morto, ossia il peso dei 4/4, si ottiene sottraendo dal peso vivo tutte le parti che non costituiscono la carne cioè ( sangue, testa, pelle, interiora, frattaglie, arti, etc.) per cui possiamo esprimere la resa al macello con la seguente espressione:
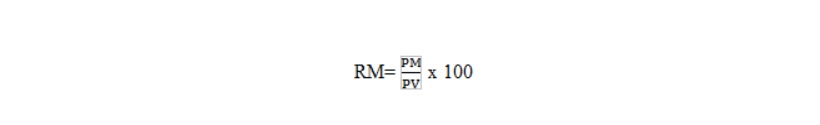 Si considerano buone rese al macello quelle > 60% del PV; nella specie bovina la resa al macello mediamente è del 55 – 60 % , danno alte rese, talvolta superiore al 70%, i vitelli di razza Charolaise, Limousine, Hereford, Aberdeen Angus. Nella specie suina i capi macellati danno rese al macello superiori dell’80%, ciò si spiega perchè in questa specie la testa e la pelle fanno parte dei 4/4; nei conigli la resa al macello va dal 45 – al 55%, negli struzzi si aggira intorno al 35 – 40 %, negli ovini e caprini la resa al macello è circa del 55 – 60 %.
Si considerano buone rese al macello quelle > 60% del PV; nella specie bovina la resa al macello mediamente è del 55 – 60 % , danno alte rese, talvolta superiore al 70%, i vitelli di razza Charolaise, Limousine, Hereford, Aberdeen Angus. Nella specie suina i capi macellati danno rese al macello superiori dell’80%, ciò si spiega perchè in questa specie la testa e la pelle fanno parte dei 4/4; nei conigli la resa al macello va dal 45 – al 55%, negli struzzi si aggira intorno al 35 – 40 %, negli ovini e caprini la resa al macello è circa del 55 – 60 %.
Neomendelismo – Altri tipi di eredità
Gli studiosi di tutto il mondo, con la riscoperta delle leggi di Mendel e nel nel rispetto delle esperienze mendeliane, fecero altri esperimenti giungendo, talvolta, a risultati differenti, onde per cui oggi si parla di altre forme di eredità.
Eredità intermedia di tipo Zea
Si caratterizza perché gli individui della F1 presentano caratteristiche intermedie tra quelle dei genitori, l’esempio è offerto incrociando mais a semi gialli con mais a semi blu, si ottengono mais a semi violetti. Negli animali incrociando polli Andalusi alcuni con piumaggio nero, altri con piume bianche chiazzate di scuro, nella F1 si ottengono polli di colore grigio azzurro. Altri esempi di eredità intermedia si ha incrociando bovini di razza Shorthorn o Durham, alcuni con mantello bianco altri con mantello rosso, si ottengono bovini a mantello bianco e rosso (ubero)
Dominanza parziale
Si verifica quando un gene, pur essendo dominante, non riesce a manifestarsi completamente nella discendenza per cui si evidenzia, anche se parzialmente, il carattere recessivo. Il fenomeno si verifica incrociando verri di razza Large White, a mantello e setole bianche, con scrofe di razza Cinta senese con mantello nero e una fascia bianca sulle spalle. I soggetti, nella 1a generazione filiale F1, nascono bianchi con macchie grigiastre (tramarchiati).
Eredità a mosaico
L’eredità a mosaico si manifesta quando in un soggetto un carattere presenta una parte di un genitore e un’altra parte di un altro genitore, esempio nell’uomo i baffi biondi e la barba nera oppure, un occhio di colore nero e l’altro di colore blu.
Eredità atavica
È il fenomeno per cui nei discendenti compaiono i caratteri non presenti nei genitori ma che esistevano negli antenati, esempio alcuni vitelli nascono con mantello fromentino, che successivamente modificano a seconda del colore di appartenenza della razza, ciò si verifica nei vitelli di razza (Chianina, Romagnola, Maremmana, Marchigiana, Piemontese, Podolica).
 Chianina con vitello (foto www.lecomunaglie.it)
Chianina con vitello (foto www.lecomunaglie.it)
Eredità ed ereditabilità
Per Eredità si intende la trasmissione dei caratteri qualitativi (caratteri a variabilità discontinua, difficilmente misurabili, influenzati da pochi geni, non influenzati dai fattori ambientali, di nessun interesse zooeconomico) da un genitore al figlio. Di contro l’ereditabilità esprime il rapporto tra ciò che un individuo riceve dai genitori (geni additivi ), ossia i caratteri quantitativi, facilmente misurabili, fortemente influenzati dai fattori ambientali, di notevole interesse economico e la variazione del carattere dovuta all’effetto ambientale. Essa è data dall’incidenza della componente additiva sulla variabilità totale di un carattere, l’ereditabilità è un concetto relativo ed è riferibile solo ai caratteri quantitativi, è l’aspetto più importante per chi si interessa di animali in produzione zootecnica. Infatti, se fosse possibile annullare l’effetto dell’ambiente sulla manifestazione dei caratteri quantitativi – rendendolo per esempio uniforme – scomparirebbe la variazione ambientale e resterebbe solo la variazione dovuta al genotipo, a questa frazione di variabilità si dà il nome di ereditabilità. Studiare l’ereditabilità di un carattere vuol dire capire quanta parte della variabilità del carattere è dovuta a fattori genetici e quanta a fattori ambientali, il parametro è indispensabile per poter applicare i programmi di selezione artificiale nel miglioramento genetico delle specie animali in produzione zootecnica.
L’ereditabilità si calcola secondo l’espresssione:
 La varianza genetica σ2G (scostamento quadratico medio dei valori rispetto alla loro media, indica il grado di variabilità dei valori, ossia l’ampiezza della dispersione dei valori rispetto alla classe fenotipica centrale) comprende quella additiva (somiglianza fra i parenti di una popolazione, dipende dai geni) su di essa si attua la selezione perché i caratteri si accumulano e possono essere trasmessi alla discendenza, quindi si può attuare il miglioramento genetico di una razza, di una popolazione o di intere famiglie,etc. e quella non additiva, è data dall’interazione dei geni nei locus, si divide di dominanza se l’interazione è tra geni dello stesso locus ed epistatica quando l’interazione è tra geni posti in loci diversi.
La varianza genetica σ2G (scostamento quadratico medio dei valori rispetto alla loro media, indica il grado di variabilità dei valori, ossia l’ampiezza della dispersione dei valori rispetto alla classe fenotipica centrale) comprende quella additiva (somiglianza fra i parenti di una popolazione, dipende dai geni) su di essa si attua la selezione perché i caratteri si accumulano e possono essere trasmessi alla discendenza, quindi si può attuare il miglioramento genetico di una razza, di una popolazione o di intere famiglie,etc. e quella non additiva, è data dall’interazione dei geni nei locus, si divide di dominanza se l’interazione è tra geni dello stesso locus ed epistatica quando l’interazione è tra geni posti in loci diversi.
La varianza non additiva non è ereditabile, come detto dipende dall’interazione dei geni, è bene non selezionare. Su soggetti parenti di una popolazione si può stimare la varianza additiva, sulle linee isogeniche e la varianza ambientale, cioè le differenze fra gli individui dovute all’effetto ambientale perciò non ereditabili né trasmissibili alla discendenza, i fattori che influiscono sulla varianza ambientale sono i fattori climatici e quelli nutrizionali, se σ2 E > σ2 G- l’ereditabilità diminuisce. Quando il valore di h2 è prossimo allo zero vuol dire che la manifestazione del carattere non dipende dalla componente ereditaria, viceversa se h2 si avvicina a 1 vuol significare che il carattere ha solo la componente genetica. In generale l’ereditabilità viene stimata in base alla somiglianza tra i genitori ed i figli, o tra i fratelli, o tra i fratellastri. Il concetto di ereditabilità non si addice al singolo animale ma ad un gruppo di soggetti allevati nello stesso ambiente (Borgioli); esempio, se per la produzione di latte si calcola h2 = 30 % se ne deduce che la superiorità o inferiorità di certe bovine è dovuta solo nel 30 % dei casi a una loro superiorità o inferiorità genetica, ma nel restante 70 % è dovuta alle diverse condizioni ambientali cui le varie bovine sono state sottoposte, il 30 % indica la percentuale di bovine che probabilmente trasmetteranno il carattere alla discendenza. Secondo il Bettini l’ereditabilità fornisce una misura della variabilità genetica di una popolazione, una misura della quantità di miglioramento ottenibile nella popolazione mediante una selezione fenotipica o massale. Ricapitolando l’ereditabilità si può calcolare col metodo del confronto dei fenotipi tra genitori e prole o tra individui con diversi gradi di parentela, infatti, se i geni sono importanti nel determinare la varianza fenotipica, allora individui imparentati dovrebbero essere più somiglianti per un certo fenotipo poiché possiedono un numero più elevato di geni in comune.

Variabilità genetica e variabilità ambientale
Come già scritto in altri capitoli i caratteri quantitativi dipendono da un grandissimo numero di geni, che si assume vengano trasmessi tramite eredità intermedia, da cui si deduce che il valore genetico o genotipico di un individuo è dato dalla somma dei geni che agiscono positivamente alla manifestazione del carattere meno la somma dei geni che agiscono negativamente sulla manifestazione dello stesso carattere (effetto genico additivo). I singoli geni del complesso poligenico ad effetto additivo che presiede alla manifestazione fenotipica dei caratteri quantitativi possono risentire anche di effetti di dominanza – recessione, cioè di una interazione tra due alleli dello stesso locus, così come di effetti di interazione tra geni posti su loci diversi (Epistasi). Oltre alla componente genetica, un ruolo importante sulla manifestazione di un carattere quantitativo è svolto dall’ambiente inteso come ambiente fisico (stabulazione, stagione, clima, età, alimentazione), che modifica in modo più o meno marcato l’espressione dei geni che controllano l’intensità di questi caratteri, da ciò si deduce che ogni carattere presenta un certo grado di variabilità dovuto all’effetto dei geni e all’effetto ambientale.
Variazioni somatiche
Sono le modifiche che interessano il soma, sono influenzate dai fattori ambientali, in primo luogo dall’alimentazione e dalla ginnastica funzionale, si differenziano, quindi, per l’ uso e il non uso di un organo (Lamarck), non interessano le cellule germinali, perciò non sono ereditarie (Darwin). Le variazioni somatiche interessano la statura, il peso, la conformazione, la pelle, il pelame, l’accrescimento, la quantità di lana etc
Variazioni genetiche o mutazioni
Le variazioni genotipiche, quelle che interessano il genoma, sono ereditarie, sorgono per effetto spontaneo o possono essere indotte dall’azione di agenti mutageni (alcool, radiazioni UV, raggi γ, raggi x, etc.) si chiamano mutazioni, queste variazioni genetiche hanno notevole importanza nella evoluzione delle specie.
La mutazione può essere genica se riguarda il singolo gene, cromosomica se interessato al cambiamento è il cromosoma, genomica se la variazione interessa tutto il cariotipo.
La mutazione può riguardare le cellule somatiche (es. tumori) in tal caso non sono ereditarie, è ereditaria la variazione del genotipo; le mutazioni, osservate per la prima volta da Darwin che le chiamò Spot, furono studiate da De Vriers, possono essere geniche, o riguardare la variazione dell’assetto cromosomico oppure, aberranti. Esempio di mutazione sono l’assenza delle corna nella razza bovina Aberden Angus, il pelo d’angora nei conigli, gli arti corti nella razza ovina Ancon, l’ipertrofia muscolare nelle razze bovine Piemontese e Charolaise. Se le mutazioni interessano le variazioni dell’assetto del numero dei cromosomi e gli effetti sono negativi si parla di aberrazioni cromosomiche come l’ idiozia mongoloide nell’uomo o trisomia del cromosoma 21.
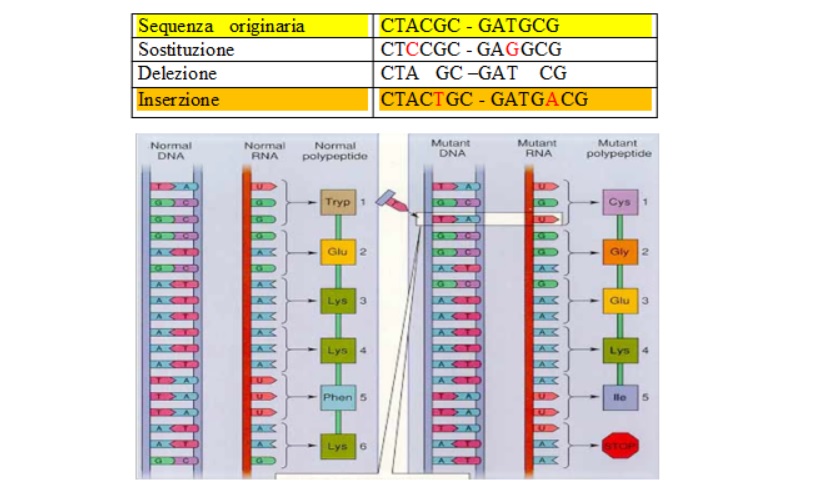
Mutazioni cromosomiche possono essere anomalie strutturali di vari tipi come appresso riportate:
Delezione: quando una parte del cromosoma è mancante.
Duplicazione: quando una parte del cromosoma è raddoppiata, causando la presenza di materiale genetico in eccesso.
Traslocazione: quando una regione di un cromosoma viene trasferita in un’altra posizione dello stesso cromosoma o di un altro; ci sono due tipi principali di traslocazioni
Inversione: quando una regione di un cromosoma inverte il suo orientamento; causando un’inversione dell’ordine dei geni.
Anello: quando le due estremità di un cromosoma si appaiano tra loro, formando un anello, quest’anomalia può comportare, o meno, perdita di materiale genetico.
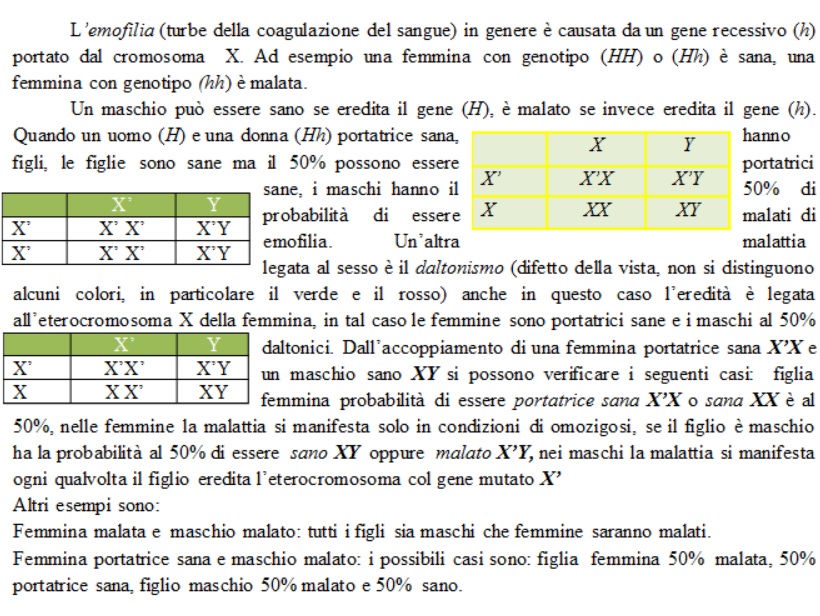
Caratteri legati al sesso
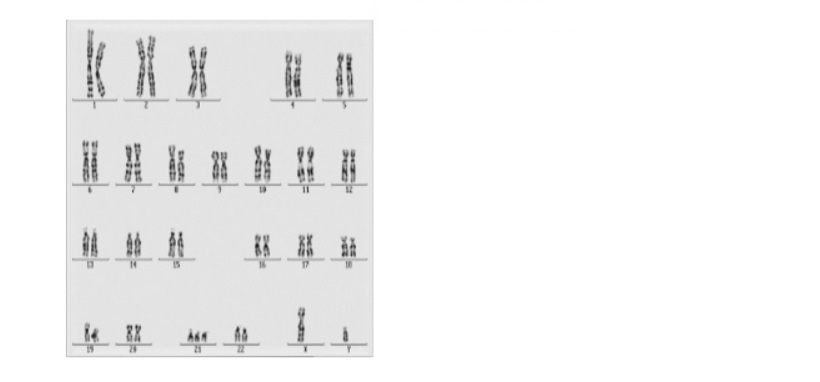
Il sesso e molti dei caratteri ad esso legati sono determinati dall’azione dei geni portati su una sola coppia di cromosomi. Nella specie umana ad esempio vi sono 22 coppie di cromosomi uguali in entrambi i sessi detti autosomi o cromosomi omologhi, ed una sola coppia di cromosomi diversi, detti perciò eterocromosomi o cromosomi sessuali indicati con le lettere X ed Y. Nella specie bovina 29 sono le coppie di cromosomi omologhi, nei suini sono 18, nella drosophila sono 3 e così di seguito. Il cromosoma Y è lungo circa 1/3 del cromosoma X e, a parte il suo ruolo nel determinare il sesso maschile, non sembra essere geneticamente molto attivo. Così, molti geni presenti sul cromosoma X non hanno una controparte sul cromosoma Y, sono questi geni legati al sesso, che vengono ereditati in modo caratteristico in alcune rare malattie che essendo legate a questo cromosoma sono dette legate al sesso, come i casi riportati.
Giuseppe Accomando, laureato in Scienze agrarie presso l’Università Federico II di Napoli, è docente di zootecnica. Curriculum vitae >>>
|




")

