
di Alessandro Cerofolini
Sistematica, origine e distribuzione
Sistematica
Il capriolo (Capreolus capreolus Linneaus, 1758) è un mammifero appartenente al superordine degli Ungulati, ordine Artiodattili, sottordine dei Ruminanti ed alla famiglia dei Cervidi.
– Classe: Mammiferi
– Sottoclasse: Uteri
– Superordine: Ungulati
– Ordine: Artiodattili
– Sottordine: Ruminanti
– Famiglia: Cervidi
– Sottofamamiglia: Capreolini
– Genere: Capreolus
– Specie: capreolus (Linneo, 1758)
La collocazione filogenetica e tassonomica del capriolo è stata in passato ed in parte lo è ancora oggi, oggetto di discussione tra i vari autori. Attualmente la maggior parte degli zoologi sostiene la tesi secondo cui il capriolo sia l’unico rappresentante euro-asiatico della sottofamiglia dei Capreolini, che differiscono dagli altri cervidi per la presenza di trofeo a punte predefinite (6). A questa sottofamiglia appartengono numerose specie del Nord e del Sud America come il cervo della Virginia (Odocoileus virginianus), il cervo delle Pampas (Ozotoceros bezoarticus), il cervo mulo (Odocoileus hemionus) e altre specie.
Recentemente la sistematica del capriolo è stata rivista, e sono sorti dubbi sulla validità di numerose sottospecie descritte in passato, ed ha inoltre riconosciuto l’esistenza di due distinte specie con i loro reciproci ecotipi: l’europea (Capreolus capreolus L., 1758) e la siberiana (Capreolus pygaus Pallas, 1771) (Danilkin, 1996; Sokolov et al. 1990). Tale tesi trova conferma dalla diversa distribuzione geografica e da differenti caratteristiche genetiche e morfometriche riscontrate tra le due specie.
Il capriolo europeo è probabilmente una specie monotipica (Danilkin, 1996; Sokolov et al. 1990) di cui in passato furono riconosciute più di venti forme descritte da Lehmann e Sagesser (1986), inizialmente classificate come sottospecie e successivamente come ecotipi. Da studi genetici e biometrici è infatti emerso che le differenze morfologiche riflettono più che una vera e propria distinzione tassonomica, adattamenti a condizioni ambientali diverse.
Anche in Italia negli ultimi anni sono state condotte indagini atte ad accertare lo status e le caratteristiche morfo-sistematiche dei rimanenti nuclei autoctoni di capriolo, riconducibili, secondo alcuni, alla sottospecie italica (Capreolus c. italicus). Questa fu descritta per la prima volta da Festa (1925), in seguito allo studio di alcune pelli provenienti dalla tenuta presidenziale di Castelporziano, nel Lazio.
Questa ipotesi fu successivamente riproposta da Lehmann (1973), il quale riconobbe in alcuni individui delle popolazioni dei monti di Orsomarso in Calabria, aspetti morfologici riconducibili alla sottospecie italica.
Grazie ad alcune indagini compiute sul DNA mitocondriale, sono state individuate nella nostra penisola due gruppi di capriolo geneticamente differenti: un gruppo presente sull’arco alpino e nell’Appennino centro-settentrionale e un altro tipico delle regioni centro-meridionali (Lovari, 2001). Quest’ultimo inoltre non sarebbe confinato soltanto nella tenuta di Castelporziano, nel Gargano e nei monti dell’ Orsomarso, come ipotizzato, ma anche in alcune aree del senese e del grossetano (Lovari, 2001).
Molto interessanti sono i dati che riguardano i caprioli del Gargano, nei quali è stato riscontrato un genotipo unico, mai riscontrato in nessuna delle popolazioni italiane analizzate. Questa caratteristica lo rende significativamente diverso sia dalle popolazioni alpine sia da quelle centro-meridionali alle quali è geneticamente più simile (Lovari, 2001).
Diventa dunque molto importante salvaguardare questi ceppi locali, inquanto dalla loro analisi genetica si potrà chiarire se almeno in quelle aree potrà essere riconosciuta la presenza di una sottospecie italiana (Lovari, 2001).
Origine
La famiglia dei Cervidi ebbe origine circa 30 milioni di anni fa nell’oligocene medio con alcune forme sprovviste di appendici frontali, canini superiori molto sviluppati e con dimensioni corporee ridotte; una specie di queste era l’ Eumeryx, la cui fisionomia ricorda da vicino i moderni mosco (Moschus moschiferus) e cervo acquatico cinese (hydropotes inermis), Cervidi dalle caratteristiche arcaiche oggi presenti nel continente asiatico (Tarello, 1991). Circa 18 milioni di anni fa, nel miocene, comparvero i primi Cervidi provvisti di appendici frontali perenni, ricoperte da pelle e prive di rosa, comunque non molto diverse dagli steli ossei dei cervidi attuali. Vere e proprie strutture caduche rinnovabili apparvero solo più tardi, circa 15 milioni di anni fa, con il genere Dicrocerus (Tarello, 1991).
Dal Miocene superiore (10 milioni di anni fa) è possibile distinguere la linea evolutiva che porterà alla specie moderna di capriolo, da quella che porta ad altri Cervidi come daino (Dama dama) e cervo nobile (Cervus elaphus) (Tarello 1991).
Derivato dalle forme del Procapreolus, il genere Capreolus ha inizio nel Pliocene circa 3 milioni di anni fa, evolvendosi progressivamente e dando origine a nuove specie simili a quella odierna. La specie Capreolus capreolus è apparsa precocemente, rispetto ad altri cervidi e bovidi, circa 700.000 anni fa (Linnel et al., 1998 Sempéré et al., 1996).
Distribuzione in Italia
Originariamente, fino alla metà del XVIII secolo il capriolo era abbondantemente diffuso in tutta l’Italia continentale e in Sicilia. Come è accaduto ad altri ungulati selvatici l’antropizzazione provocò una progressiva ma marcata diminuzione sia numerica sia di areale. Questo processo caratterizzò tutto il territorio italiano e fu particolarmente intensa nell’Italia meridionale e in Sicilia, dove il capriolo arrivò all’estinzione prima della fine del XIX secolo (Pedrotti et al., 2001).
Le principali cause che determinarono il declino della specie sono da imputare all’esasperato sfruttamento agricolo e forestale del territorio, unito all’aumento indiscriminato della pressione venatoria. I motivi scatenanti furono diversi da regione a regione, infatti, nell’Italia centro-settentrionale vi fu l’intenso disboscamento di collina e montagna (soprattutto nel settentrione) e l’incontrollata attività venatoria (praticata con il cane da seguito nel Nord e nell’appennino Tosco-Emiliano), nell’Italia meridionale fu soprattutto l’espansione delle attività agro-silvo-pastorali (Tarello, 1991). La fase più critica di questo fenomeno corrisponde al periodo immediatamente successivi alla seconda guerra mondiale: la specie arrivò a toccare il suo minimo storico nel 1945, infatti veniva stimata una presenza complessiva di non più di 10.000 capi, distribuiti tra Valtellina (Lombardia), parte delle Alpi centro-orientali (Trento, Bolzano, Belluno e Udine) e in Italia centrale (Maremma toscana); alcuni nuclei isolati si conservarono anche nel Gargano ed in Calabria.
A partire dalla fine degli anni ’60 invece si ha una netta inversione di tendenza, infatti la specie appare in attiva espansione tanto da rioccupare una parte considerevole del proprio areale storico (Pedrotti et al., 2001). Questo processo fu reso possibile da una serie di eventi concomitanti: l’abbandono delle tradizionali attività rurali in vaste zone montane e di alta collina, con il conseguente ripristino della vegetazione naturale che ha portato a una maggiore disponibilità di habitat idonei, l’introduzione di norme tese a regolamentare o vietare la caccia alla specie, l’immigrazione in nuovi territori di soggetti provenienti da nuclei residui, spesso integrata da operazioni di reintroduzione con individui di provenienza alpina o centro-europea.
Queste ultime operazioni sebbene abbiano contribuito in maniera importante al recupero della specie, hanno causato l’inquinamento genetico delle popolazioni autoctone interessate dal fenomeno (Casanova et al., 1993). Dopo una prima fase dove vi fu un accrescimento demografico di tipo “irruttivo”, cioè esplosivo, il capriolo si è assestato su gran parte dell’area montana, dal piano basale al limite superiore della vegetazione arborea e arbustiva, di contro, come avviene negli altri paesi europei, manca quasi completamente dalle pianure intensamente coltivate. Attualmente in Italia possiamo distinguere due grossi subareali: il primo comprende senza soluzione di continuità tutto l’arco alpino, l’Appennino ligure e lombardo sino alle province di Genova e Pavia; il secondo si estende lungo la dorsale appenninica, dalle province di Parma e Massa Carrara sino a quelle di Terni e Macerata, occupando anche i rilievi minori che formano l’Antiappennino toscano delle province di Pisa, Siena, Livorno, Grosseto e la Maremma Toscana. Piccoli areali disgiunti si trovano nell’Italia centro-meridionale, in cui si trovano nuclei isolati, resti delle popolazioni anticamente presenti nella penisola (Castelporziano, Orsomarso, Gargano) o frutto di più o meno recenti immissioni (Parco nazionale d’Abruzzo, Sila).
Le attuali popolazioni di capriolo diffuse sull’arco alpino e appenninico, per le origini descritte, debbono essere attribuite alla forma tassonomica C. c. capreolus, mentre le uniche popolazioni riconducibili alla forma C. c. italicus, sarebbero presenti nella tenuta presidenziale di Castelporziano (Roma), nella foresta Umbra (Gargano, Foggia), nei monti dell’Orsomarso (Cosenza) ed in alcune aree del senese e del grossetano (Lovari, 2001). La consistenza effettiva sul territorio nazionale dovrebbe aggirarsi introno ai 400.000 capi, anche se una stima precisa risulta difficile a causa delle intrinseche difficoltà di censimento legate alle caratteristiche biologiche ed ecologiche della specie (Pedrotti et al., 2001); inoltre i censimenti vengono realizzati solamente nelle aree in cui viene esercitata l’attività venatoria, con difformità notevoli delle tecniche di conteggio utilizzate.
La specie può essere considerata ampiamente distribuita e relativamente comune in Italia settentrionale e centrale, in cui il capriolo occupa quasi l’80% del suo areale potenziale, pur con qualche differenza strettamente legata alla qualità della gestione venatoria di ciascuna regione o provincia. Infatti la consistenza delle popolazioni risulta massima nell’Appennino centro-settentrionale ed elevata nell’arco alpino centro-orientale, dove viene riscontrato uno stato di conservazione soddisfacente ed in progressivo miglioramento; mentre diminuisce sul versante alpino occidentale, a causa di alcuni fattori limitanti ancora presenti. Per quanto riguarda l’Italia centro-meridionale lo stato di conservazione del capriolo risulta ancora estremamente precario, la distribuzione è frammentaria e la percentuale di occupazione dell’areale potenziale non supera l’ 1%.
Risulta quindi di primaria importanza in questo contesto la messa in atto di azione volte a tutelare dall’estinzione e dall’inquinamento genetico i nuclei autoctono residui, favorendone la diffusione e riducendo in maniera decisa il bracconaggio ed il randagismo canino, che appaiono ad oggi come i principali fattori limitanti nel meridione (Pedrotti et al., 2001). Con queste premesse ci possiamo aspettare per il futuro un ulteriore espansione dell’areale, specialmente per il centro-sud, dove esistono ancora molti spazi idonei alla colonizzazione.
Origine e distribuzione in Toscana
In Toscana, come in molte altre zone d’Italia, il capriolo è stato sottoposto ad intensa gestione, che ne ha causato frequenti estinzioni locali e conseguenti reintroduzioni. Operazioni di questo tipo furono condotte dal Corpo Forestale dello Stato durante gli anni ’50 nelle foreste demaniali dell’alto pistoiese, dove secondo Mattioli (1994) furono liberati 23 caprili di origine alpina, fra il 1968 e il 1973 nelle foreste dell’alto Serchio, nelle riserve naturali di Lamarossa, dell’Orecchiella e di Pania di Corfino, con esemplari provenienti sia dalla Maremma che dal Trentino (Masseti, 2003). Le popolazioni attualmente presenti in provincia di Arezzo sarebbero in parte derivate da ripopolamenti effettuati con alcuni esemplari di provenienza alpina durante gli anni ’30 e ’50, in parte avrebbero mantenuto la loro radice autoctona in quanto discendenti da animali presenti originariamente nelle Foreste Casentinesi (Lorenzini et al., 1996). Infatti qui si hanno testimonianze storiche che confermano la sicura autoctonia e presenza della specie durante il 1800: a differenza di ciò che accadde per gli altri ungulati (cervo, daino, muflone) per il capriolo non avvennero immissioni nel corso del XIX secolo, sia per la difficoltà nel reperire il piccolo cervide, sia per la probabile presenza in zona di alcuni esemplari. Soltanto nel 1933 furono immessi alcuni esemplari di capriolo a cura dell’ Azienda di Stato delle Foreste Demaniali (A.S.F.D), sebbene è noto che alcuni di essi morirono in tempi brevi a causa del bracconaggio.
Durante la seconda guerra mondiale e nei periodi immediatamente successivi, le popolazioni di capriolo subirono un drastico calo numerico (sembra che nel 1946 fossero rimasti solo una trentina di esemplari), sebbene da fonti note è possibile rilevare che esiste una continuità genetica tra le popolazioni del periodo prebellico e le popolazioni attuali (Mazzarone et al., 2000). Si ritiene inoltre che i caprioli presenti oggi nel territorio senese avrebbero avuto origine da immissioni effettuate a partire dagli inizi del ‘900 con animali provenienti sia dalla Maremma laziale, che con soggetti alloctoni provenienti dalla Gran Bretagna e dalla Cecoslovacchia (Lorenzini at al., 2000).
Possiamo quindi dedurre che l’incremento numerico della specie e il conseguente ampliamento dell’areale a cui si è assistito negli ultimi decenni nel territorio regionale sia da imputare principalmente alle numerose reintroduzioni, ed in misura minore alla diffusione naturale delle restanti popolazioni indigene, che si ritrovano oggi solo in alcune aree della Maremma e del senese.
Infatti, l’attuale espansione sia naturale che artificiale degli ecotipi autoctoni e di quelli alloctoni ha portato ormai ad una sovrapposizione degli areali, per cui sembra piuttosto difficile riuscire in futuro a preservare inalterate le caratteristiche genetiche di uno specifico capriolo dell’Italia centro-meridionale (Masseti et al., 2001).
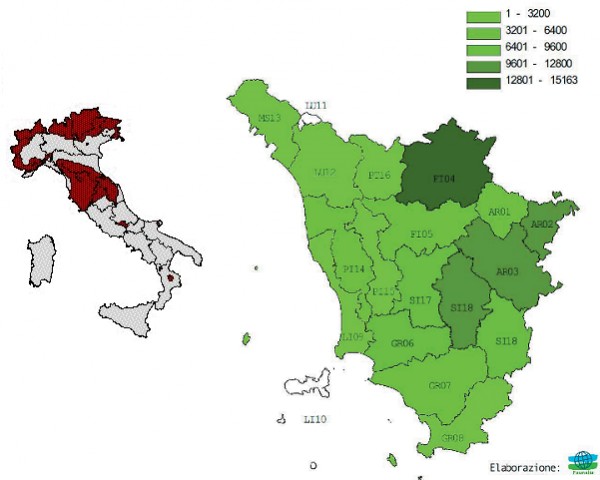
La Toscana appare la regione italiana che ospita le popolazioni di capriolo più numerose, in cui si registra una presenza stimata nel 1999-2000 di 107.300 capi, distribuiti su gran parte del territorio regionale (Pedrotti, 2001).
Il Piano faunistico regionale della Toscana 2007-2010 riporta nella Tabella 34: Distretti per la caccia al capriolo per l’anno 2004, 100.477 capi derivanti dalla somma dei capi stimati nei singoli ATC. La distribuzione sul territorio regionale è quella riportata in figura sottostante.
La specie è diffusa in tutto l’Appennino tosco-emiliano, dalla Lunigiana fino a Bocca Trabaria, comprendendo la Garfagnana, il pistoiese, il Mugello, il Pratomagno ed il Casentino e la si ritrova uniformemente diffusa anche a sud dell’Arno fino al confine meridionale della Toscana, essendo anche presente in provincia di Lucca, di Livorno, di Siena e di Grosseto (Masseti et al., 2001).
Distribuzione del Capriolo in Italia (al 2005) e in Toscana
Caratteristiche morfologiche
Il capriolo è il più piccolo cervide della fauna europea, molti profani lo confondono spesso con i piccoli di cervo nonostante siano individui con notevoli differenze morfologiche. Il capriolo ha una conformazione fisica slanciata, corpo raccolto, la groppa più alta del garrese (Lister et al., 1998) fa si che il treno posteriore sia più robusto di quello anteriore: tipica struttura dei saltatori. La testa è corta, il profilo appare nettamente triangolare (nel maschio più che nelle femmine),terminante anteriormente in un muso minuto, stretto e nudo. Le orecchie e gli occhi sono grandi, con le prime che raggiungono i 2/3 in lunghezza della testa. La coda è praticamente invisibile.
Dimensione e sviluppo corporeo
Le differenze tra i due sessi per quanto riguarda i valori dei principali parametri morfometrici sono praticamente irrilevanti, evidenziando così uno scarso dimorfismo sessuale, anche se in genere le femmine presentano dimensioni di poco inferiori al maschio (Lister et al., 1998).
Un individuo di dimensioni medie è alto al garrese circa 60-75 cm, mentre la lunghezza totale varia da 1 a 1,20 m (Il cacciatore di selezione, Casanova 2006). Il peso è da mettere in relazione all’ambiente e quindi all’offerta alimentare, generalmente in un individuo sano e ben sviluppato dovrebbe essere compreso tra i 20 e i 30 kg, mentre certi individui che abitano in zone particolarmente favorevoli possono arrivare anche fino a 36 kg; Le femmine sono in media più leggere dei maschi di 2 o 3 kg. Alla nascita il capriolo pesa da 0,9 a 1,8 kg circa ed è caratterizzato da uno sviluppo ponderale molto rapido: a due settimane di vita raddoppia il proprio peso, mentre a 8 mesi raggiunge già il 70% del peso definitivo (Andersen et al., 1998; Mustoni et al., 2002; Sempéré et al., 1996), che in caso di sviluppo normale viene raggiunto nel quarto anno di vita nei maschi e nel terzo anno nelle femmine. Nel corso della senescenza si può assistere ad un regresso di peso, maggiormente visibile nei maschi.
La variabilità di peso e dimensioni presente all’interno della specie è condizionata da fattori individuali (peso e condizione fisica della madre, origini genetiche), sociali (densità di popolazione) e ambientali (altitudine, clima, disponibilità trofiche sia in termini qualitativi che quantitativi) (Andrersen et al., 1998; Mustoni et al., 2002; Toso et al., 1998). I caprioli però non pesano ugualmente durante il corso dell’anno, questo fenomeno si riscontra maggiormente nei maschi dove le variazioni di peso possono essere notevoli. Nei mesi di maggio, giugno e ottobre è raggiunto il massimo della mole, mentre durante il periodo degli amori gli individui pesano sensibilmente meno. Una continua diminuzione di peso avviene pure nei mesi invernali quando il cibo scarseggia, per le femmine vale lo stesso ciclo essendo impegnate in estate nell’allattamento della prole.
Mantello
Come tutti i cervidi il mantello dei piccoli si presenta di colore bruno scuro e picchiettato lungo la schiena e i fianchi (Lister et al., 1998), nell’arco di circa due mesi tali macchie tendono a scomparire per lasciare spazio al mantello estivo rossiccio, analogo a quello dell’adulto (Mustoni et al., 2002). Il mantello subisce due mute annuali: la muta primaverile fa assumere una viva colorazione bruno-rossastra, sebbene possano essere osservate variazioni di tonalità in base all’età degli individui. Spesso gli animali più giovani e sani hanno il mantello estivo più vivo, mentre vecchi e malati sono più chiari. In ottobre-novembre il pelo è molto più fitto e lungo, con una colorazione grigio-brunastra che ben mette in risalto la zona posteriore completamente bianca denominata “specchio anale”. Questa zona è un utile strumento per riconoscere i sessi poiché presenta una forma a rene o fagiolo nel maschio e a cuore rovesciato nella femmina; in quest’ultima inoltre è possibile distinguere una “falsa coda”, ovvero un ciuffo di peli color crema che protegge l’organo genitale.
La muta primaverile avviene tra aprile e giugno ed è un fenomeno piuttosto vistoso che può durare anche una ventina di giorni, caratterizzato da una massiccia caduta di ciocche del mantello invernale; mentre la muta autunnale, che si manifesta tra settembre e ottobre (raramente fino a novembre), risulta meno appariscente e avviene in tempi piuttosto rapidi. Da rilevare come il periodo delle mute vari in relazione dell’età e allo stato di salute dell’individuo: il mantello infatti viene cambiato prima dai giovani, seguiti poi dai subadulti, dagli adulti, dagli individui malati o deboli, dai vecchi (Mustoni et al., 2002; Nicoloso e Orlandi, 2002). Caratteristica peculiare del capriolo che lo distingue dagli altri Cervidi italiani, è quella di avere il labbro inferiore completamente bianco.
Organi sensoriali e ghiandole
Particolarmente sviluppati risultano nel capriolo i sensi dell’olfatto e dell’udito. L’importanza di quest’ultimo è evidenziata dalla forma e dalle dimensioni delle orecchie, nonché dalla loro estrema mobilità (Sempéré et al., 1996; Tarello, 1991). L’olfatto ha un grande rilievo nell’ambito dei contatti e delle relazioni sociali, in particolar modo durante la fase territoriale e degli accoppiamenti. Nei maschi infatti la regione frontale è cosparsa di numerose ghiandole che in primavera-estate emettono una sostanza sebacea odorosa, che serve per marcare il territorio. Anche nella zona metatarsale e interdigitale di ambedue i sessi sono presenti ghiandole odorifere, attive tutto l’anno, le quali sembrano assumere funzione di richiamo “amoroso” nel primo caso e di riconoscimento individuale nel secondo (Sempéré et al., 1996; Liberg et al., 1998).

Maschio e femmina di capriolo – Maschio che marca il territorio
La vista è il senso meno sviluppato, la posizione fortemente laterale degli occhi rende quasi impossibile la visione binoculare, e sebbene i caprioli siano molto capaci nell’individuare ogni piccolo movimento, hanno una scarsa capacità a definire un oggetto immobile (Tarello, 1991).
Palco
Nel capriolo, come in tutti gli altri cervidi ad eccezione della Renna, solo il maschio possiede appendici frontali. I palchi sono strutture caduche formati da un vero e proprio tessuto osseo, soggetti ad un ciclo annuale. La loro composizione è prevalentemente di carbonato di calcio. I palchi di un individuo adulto sono costituiti da due aste, denominate stanghe, con tre diramazioni dette cime o punte che dalla anteriore alla posteriore prendono il nome di “oculare”, “vertice” e “stocco”. La parte basale, sporgente sopra la pelle del cranio, si presenta ingrossata a forma di corona ed è chiamata “rosa”. Sulle stanghe sono presenti, in particolar modo sulla parte interna e basale, delle escrescenze a forma di goccia chiamate “perle”.
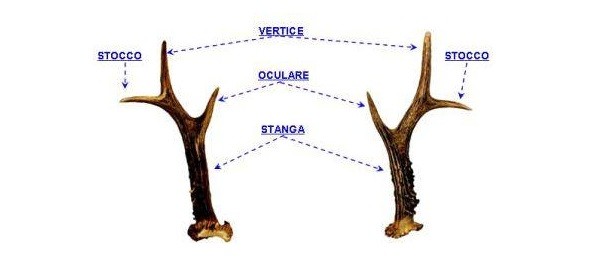
Palco del capriolo e relativa terminologia.
La formazione dei palchi inizia per la moltiplicazione e la modificazione di particolari cellule poste all’estremità delle ossa frontali del cranio grazie alle quali si ha la produzione di tessuto osseo. Durante la formazione il palco è rivestito da una cute pelosa ricca di vasi sanguigni e terminazioni nervose chiamata “velluto”. In questo periodo infatti i palchi sono molto sensibili provocando dolore all’animale se accidentalmente urtati; con l’avanzare dei processi di ossificazione i vasi sanguigni del velluto vengono occlusi provocandone così la morte, anche l’aspetto diverso assunto dalle cime denota la necrosi del velluto che non è più globoso ma piatto e di colorito più smorto. Il capriolo si libera di questo tessuto ormai inutile, costituito da pelle attaccata alle stanghe, mediante un operazione detta di “pulitura” ottenuta tramite lo sfregamento contro tronchi o arbusti. Tutti i processi descritti sono regolati da ormoni, in particolare la somatropina (ormone della crescita la cui concentrazione è massima in primavera) ed il testosterone (ormone maschile, responsabile dell’ossificazione del palco). Dopo 80-120 giorni il periodo degli amori i palchi cadono: ciò avviene per l’azione di cellule poste all’estremità delle prominenze frontali le quali hanno sempre conservato la loro struttura ossea viva, in conseguenza di tale attività si formano delle zone di riassorbimento del tessuto osseo che determinano una superfice di distacco; appena i palchi sono stati deposti, questa linea di frattura cicatrizza e inizia subito il processo di formazione del nuovo trofeo (Casanova et al., 1993).
I palchi spuntano per la prima volta l’autunno successivo alla nascita. Nel giovane maschio la formazione inizia a 5-6 mesi e continua lentamente fino a dicembre-gennaio concludendosi con la formazione di due piccole prominenze frontali dette “bottoni”; dopo poche settimane dal termine del loro sviluppo, i piccoli palchi vengono persi. Segue la formazione, molto rapida, dei primi e veri propri palchi. La ricrescita si ha in febbraio-marzo, essa è in relazione sia con l’anzianità (avviene prima negli individui vecchi e adulti), sia con il buono stato di salute dell’animale, sia a circostanze climatiche locali non sempre perfettamente valutabili. Il capriolo maschio di un anno compiuto ha due semplici aste e viene detto “puntuto”. L’anno successivo i palchi presentano due punte con aspetto di piccole forcelle “forcuto” ed infine l’anno successivo compiuto il terzo anno di età, il trofeo del capriolo è completamente sviluppato con tre punte per palco “palcuto” (Nicoloso e Orlandi, 2002; Perco 1995).
La deposizione avviene dalla fine di ottobre a tutto novembre-dicembre ed anche in questo caso gli adulti precedono i giovani. Per quanto riguarda le dimensioni, la lunghezza delle stanghe va dai 20 (18) ai 25 (30) cm (Lister et al., 1998; Perco, 1995; Sempéré et al., 1996). Le cime, spesso diseguali, variano dai 2-3 cm ai 5 (8) cm. Peso e volume variano invece molto. Il peso si aggira introno ai 180 e i 250 gr (Lister et al., 1998; Perco, 1995; Sempéré et al., 1996) . Orami è accertato come i palchi svolgano una funzione determinante nelle relazioni sociali in quanto esprimono il vigore fisico del maschio definendone il rango sociale. Quindi, se il regime alimentare è buono, si possono (in casi rari) avere soggetti forcuti ad un anno e palcuti a due, importante è la qualità degli alimenti che devono avere un alto contenuto di calcio e fosforo, necessari per una completa ossificazione. La denutrizione, infatti, porta alla formazione di palchi deboli, poco perlati, con cime piuttosto corte. Da ricordare come l’ossificazione sia regolata dall’ormone testosterone, prodotto dai testicoli, quindi lesioni o malattie ad essi possono portare ai cosiddetti “palchi a parrucca” nei quali si ha un accrescimento continuo senza solidificazione del tessuto osseo. Il materiale ricade fino a coprire tutto il muso determinando così cecità. Un buono sviluppo dipende anche dallo stato psichico del soggetto; fattori di stress che ne possono turbare l’equilibrio sono: densità eccessive, condizioni di forte competizione con altre specie animali, influenze dell’uomo, disequilibrio dei ranghi sociali. Possiamo quindi affermare che dall’osservazione attenta dei palchi possiamo trarre importanti informazioni sulla salute della popolazione.
Dentatura
Il capriolo è caratterizzato dall’assenza degli incisivi e dei canini superiori, al loro posto vi è infatti un ispessimento della mucosa orale chiamato “placca dentale”. Caratteristico risulta quindi il “morso”, la superficie dei rametti brucati appare sfilacciata anziché tagliata di netto. La dentatura da latte, presente alla nascita, è composta da un totale di 20 denti. La sostituzione della dentatura da latte con quella dell’adulto, si completa normalmente entro il 12° mese di vita (Aitken, 1975) e comprende un totale di 32 denti.
I canini mascellari raramente sono presenti, ma possono raramente ritrovarsi in entrambi i sessi, anche se risultano meno frequenti nelle femmine, questo a supporto della tesi che in un lontano passato i maschi presentavano canini molto sviluppati come carattere secondario maschile. Nella mandibola i canini hanno ormai assunto una morfologia ed una funzionalità da incisivi. L’esame della mandibola consente, con una certa approssimazione, la stima dell’età dell’animale. Una prima osservazione consente infatti di distinguere i soggetti inferiori all’anno di età da quelli che l’anno già superato: gli elementi caratteristici di questo passaggio sono la sostituzione del 3° premolare, che dalla forma tricuspide (tre punte, “p3”) passa a quella bicuspide (due punte “P3”) e la completa eruzione del 3° molare M3 (Nicoloso e Orlandi, 2002).
Dopo i 12 mesi la stima dell’età del capriolo può essere fatta solo attraverso l’analisi del grado di usura della tavola masticatoria a livello di molari e premolari, ovviamente questo metodo risulta abbastanza approssimativo in quanto risente di vari fattori caratteristici dell’individuo come la mineralizzazione, la meccanica masticatoria e dell’ambiente in cui vive come clima ed alimentazione.
Eco-Etologia
Alimentazione
Il capriolo, a causa del suo elevato metabolismo e della sua scarsa capacitò ruminale, pari a circa il 6-8,5% del peso corporeo (Sempéré et al., 1996; Tarello, 1991) , necessita di un’alimentazione selettiva che privilegia frazioni vegetali povere di fibra e facilmente digeribili, ad alto contenuto energetico e proteico. La digestione è decisamente rapida ed il ritmo giornaliero è suddiviso in numerosi periodi di alimentazione (dai 10-11 in primavera-estate ai 6-7 durante l’inverno), che sono più frequenti durante le ore crepuscolari (alba e tramonto), sebbene tale attività possa essere intensa durante tutto l’arco della giornata (Tarello, 1991).
Il capriolo, eccelso selettore, non è da ritenersi un esclusivo consumatore di alberi e arbusti come descritto in passato, poiché alcune indagini hanno dimostrato che il suo nutrimento si basa soprattutto su erbe ad alto contenuto proteico (leguminose foraggere), mentre i germogli delle varie latifoglie assumono un ruolo preminente solo quando non vi è adeguato pascolo erbaceo (Casanova, 1990). Le parti legnose delle piante sono utilizzate solo raramente (Duncan et al., 1998). Mediamente la dieta del capriolo è costituita da specie arboree e arbustive per il 25%, da piante erbacee dicotiledoni (leguminose) per il 54%, e monocotiledoni (graminacee) per il restante 16% (Sempéré et al., 1998). La specie comunque presenta ampie variazioni nella quantità e nella proporzione di essenze vegetali consumate, in relazione all’offerta alimentare e ai diversi momenti del ciclo biologico in cui si trova. La maggior disponibilità di alimento che il territorio possa offrire si ha in giugno-luglio, periodo che coincide con l’aumento delle richieste energetiche da parte dell’animale a causa dell’allattamento, dello sviluppo dei piccoli e del rituale che accompagna la riproduzione (Perco, 1995; Sempéré et al., 1996). Durante l’inverno il livello del metabolismo basale diminuisce, adattandosi alla penuria invernale, in questo periodo i caprioli snobbano campi e prati, il loro nutrimento è infatti quasi interamente consumato nel bosco e riguarda germogli di rovo o lampone, ghiande, castagne, faggiole e funghi; con l’avanzare dell’inverno il cibo è costituito soprattutto dalle cime di querce, di abete bianco (ove presente) e in genere di tutti gli alberi, arbusti e cespugli (Perco, 1995). Verso la fine della stagione invernale, che solitamente rappresenta il periodo critico dal punto di vista alimentare, il capriolo si rivolge in gran parte agli apici vegetativi della rinnovazione forestale (Casanova, 1990).
Organizzazione sociale
Il capriolo è un animale tipicamente territoriale, manifesta cioè un comportamento stagionale attraverso cui i soggetti adulti di rango più elevato entrano in possesso di una determinata area, ritenuta la più favorevole, all’interno della quale non viene tollerata la presenza di altri individui dello stesso sesso (Hewison et al., 1998). La territorialità non è prerogativa dei solo maschi, anche le femmine durante il periodo dei parti e nelle fasi immediatamente successive individuano e difendono un loro territorio (Hewison et al., 1998). L’indole territoriale del capriolo ne spiega in parte anche la socialità: escluse forti relazioni parentali che si formano tra madre e prole, il capriolo non forma veri e propri branchi, limitandosi a qualche momento di apparente aggregazione quando le circostanze lo rendono necessario, ad esempio in caso di carestie invernali. Si parla quindi di un organizzazione sociale basata sul gruppo familiare invernale tipico, costituito da 3-5 individui con organizzazione matriarcale: alla madre infatti spettano i compiti di scelta di località di pascolo, di guida negli spostamenti, di decisione se prendere la fuga o rimanere (Casanova et al., 1993; Perco, 1995). Le unità familiari sono in genere stabili e composte dalla femmina adulta capogruppo con i suoi piccoli dell’anno, seguita eventualmente dalla figlia sottile e dal maschio che l’accompagna (Hewison et al., 1998; Perco, 1995).
Fasi annuali del ciclo biologico
All’interno della popolazione, le attività e le interazioni fra i vari individui non sono costanti, ma variano nel corso dell’anno, influenzate dal mutare delle stagioni. Il comportamento del capriolo può essere suddiviso in varie fasi, che costituiscono i momenti di maggiore attività sociale della specie. A partire dai tre anni di età, alla fine dell’inverno, i maschi tentano di delimitare un territorio, all’interno del quale nessun altro individuo di esso maschile è ammesso. I rapporti tra i vari individui, all’interno di una popolazione, non sono costanti ma variano nel corso dell’anno, il comportamento dei caprioli, infatti, può essere così diviso in varie fasi che costituiscono i momenti di maggiore attività sociale. Tali periodi corrispondono alla fase gerarchica, alla fase territoriale ed al periodo degli amori. Verso la fine dell’inverno, i maschi più vecchi e più forti hanno un trofeo già quasi formato, ben visibile, anche se talvolta è ancora in velluto o sono da completare i processi di solidificazione. Il maschio che lo porta tende già ad assumere atteggiamenti di intolleranza verso gli altri maschi più giovani o deboli, verso la metà di Marzo, i processi di formazione dei palchi sono terminati e quindi possono essere usati come arma di offesa: inizia così la fase gerarchica. A questo proposito si possono distinguere alcuni tipi di atteggiamenti, riassumibili in tre categorie: – Comportamento di imposizione, – di sottomissione, – di minaccia.
Il comportamento tipico di imposizione ha soprattutto la funzione di mostrare la propria forza e quindi di impressionare o intimidire i rivali. Molto raramente si può arrivare ad uno scontro diretto, in quanto potrebbe essere dannoso per entrambi i contendenti. I due maschi si posizionano uno di fronte all’altro, con il collo ben dritto, la testa piegata di fianco e le orecchie rivolte all’indietro. Se il rivale appartiene ad un rango sociale inferiore riconoscendo la propria inferiorità fisica, mostrerà un comportamento di sottomissione, tendendo a “farsi piccolo”, assumendo una posizione raccolta con il collo tenuto orizzontale. Il comportamento di minaccia è invece un insieme di atteggiamenti tesi a dissuadere, mediante intimidazione e aggressione; il capriolo punta il muso verso terra, avanza verso l’avversario tenendo le orecchie ben dritte, tutto questo può essere rafforzato anche dalla “raspatura”. Il rango sociale viene così a fissarsi tra i maschi di diversa forza ed età mediante imposizione, minaccia, fughe determinate alcune volte da brevi inseguimenti. Se dopo i vari atteggiamenti di imposizione, nessuno dei due contendenti si allontana o si sottomette, si arriva alla minaccia: i maschi si avvicinano e, se ancora nessuno dei due scappa, inizia la lotta. Normalmente i combattimenti non sono violenti, e chi riconosce la propria inferiorità, dopo i primi attacchi è costretto a fuggire. I maschi più vecchi, cioè quelli che hanno un processo di ossificazione del palco precoce rispetto agli altri, sono solitamente i più alti nella scala gerarchica della popolazione.
Alla fine della primavera tutti i maschi sono venuti a “conoscenza” dei loro antagonisti e tutti hanno un ruolo ben definito all’interno della popolazione. In questa fase quindi ogni maschio sa qual è la sua posizione. Alla fase gerarchica segue la fase territoriale, l’inizio di tale fase non è esattamente determinabile, essendo comportamenti che spesso sfumano gli uni negli altri. Il territorio ha dei confini tracciati tramite vari segnali, che possono essere visivi (fregoni e raspate), acustici (abbaio) ed olfattivi ed è difeso dal maschio che lo possiede. Solo i maschi di rango ampiamente inferiore sono ammessi e solo in atteggiamenti di totale sottomissione, le femmine sono naturalmente ben accette. L’ampiezza del territorio è variabile, in media è fra i 10 e i 20 ha per i territori migliori, a fra i 20 e i 30 ha per i peggiori, questo perché generalmente i più piccoli sono più ricchi di alimenti, più sicuri e meglio difendibili. Il territorialismo maschile risulta importante anche perché consente di regolare la densità della popolazione determinando uno sfruttamento più razionale dell’ambiente.
La stagione degli amori si concentra tra la metà di luglio e la metà di agosto e l’accoppiamento avviene all’interno del territorio del maschio, in quanto le femmine in questo periodo hanno solitamente un home range coincidente con esso. Infatti il territorio di un maschio può coincidere parzialmente o totalmente con quello di una o più femmine, di norma tre, in casi eccezionali anche dieci; ciò sarebbe anche all’origine dell’alta incidenza di accoppiamenti tra soggetti consanguinei che si riscontra all’interno delle popolazioni di capriolo, sebbene tale fenomeno sia leggermente ridotto dalle brevi escursioni dei maschi fuori dal loro territorio alla ricerca di altre femmine. Il capriolo è quindi una specie poligama, in quanto nella stessa stagione riproduttiva è in grado di accoppiarsi con una o più femmine, sebbene non formi un harem. Infatti, a differenza di altri cervidi, il capriolo forma coppie più o meno stabili, di natura diversa a seconda che si tratti di una femmina adulta o di una giovane. L’unione tra maschi e femmine giovani generalmente ha una durata maggiore (la coppia può formarsi già nel periodo invernale e perdurare anche dopo l’accoppiamento) rispetto alle coppie formate con le femmine adulte, ancora impegnate nelle cure parentali. Le femmine di capriolo sono monoestrali e la durata dell’estro è di circa 36-48 ore. Il calore si manifesta con un picco tra la fine di luglio e l’inizio di agosto, da prima nelle sottili e dopo nelle adulte. In queste ultime avviene mediamente a 68 giorni dal parto, in quanto sono ancora impegnate nella cura della prole, e il manifestarsi del calore è negativamente correlato con le frequenti poppate. La fine del periodo degli amori segna un momento di grandi modificazioni nella socialità del capriolo: le madri riacquistano il rapporto con la prole, determinando così un assestamento definitivo bel gruppo madre-figli, questa fase coincide con il periodo tardo autunnale e invernale in cui si viene a ristabilire il gruppo familiare tipico.
In condizioni particolarmente rigide (come innevamento forte e prolungato), più gruppi familiari possono concentrarsi nelle poche aree di pascolo che offrono maggiori risorse trofiche, per dare così origine ai cosiddetti “assembramenti”, pseudo-branchi di scarsa coesione che si sciolgono al cessare delle condizioni sfavorevoli ed in cui ogni gruppo mantiene la propria individualità. Con l’approssimarsi del parto (maggio-giugno) avviene la completa disgregazione del gruppo invernale, in quanto la femmina adulta si isola per partorire i piccoli, generalmente due, anche se non sono rari i parti trigemini. La gravidanza del capriolo è tipica, infatti è possibile distinguere una diapausa embrionale di circa 4-5 mesi, a cui fa seguito la gravidanza vera e propria, anch’essa di circa 5 mesi: in totale quindi la gestazione completa dura dai 285 (264) ai 300 (318) giorni (Sempéré et al., 1996; Tarello, 1991). Nelle prime settimane di vita i caprioletti sono ancora molto immaturi e trascorrono poco tempo insieme alla madre. Questa infatti si allontana per alimentarsi, rimanendo comunque nei dintorni e ritornando dalla prole per allattarla più volte al giorno, seppur per brevi periodi. I piccoli rimangono apparentemente senza difesa: in realtà la loro immobilità, il mantello mimetico e l’assenza di odore caratteristico sono una garanzia di salvezza nei confronti dei predatori, diventando però una delle più alte cause di mortalità quando i piccoli si trovano in prati che vengono falciati con mezzi meccanici, che finiscono per ucciderli (Perco, 1995; Sempéré et al., 1996). Una peculiarità del capriolo è la lunghezza del “periodo critico dell’imprinting”, che si verifica nella terza-quarta settimana di vita fino a fine luglio, cioè l’evoluzione del rapporto a distanza in quello di contatto, durante il quale il piccolo, secondo un meccanismo innato inizia ad apprendere, a riconoscere i membri della propria famiglia e della propria specie, a discriminare gli estranei e i potenziali pericoli, in modo che l’attaccamento alla madre diventa stabile ed irreversibile (Perco, 1995).
Habitat ed uso dello spazio
L’optimum ecologico del capriolo è rappresentato da territori di pianura, collina e media montagna, nei quali si sviluppa un mosaico ad elevato indice ecotonale (Perco, 1995; Sempéré et al., 1996). L’ambiente più adatto alla specie originariamente era la foresta di latifoglie mesofite con folto sottobosco, e la stessa macchia mediterranea, ambedue inframezzate da radure, pascoli e colture foraggere. Sono da ritenersi quindi ambienti buoni per il capriolo i querco-carpineti, i querco-castagneti ed i querceti dei boschi planiziali, mentre la faggeta risulta meno adatta perché priva quasi ovunque di sottobosco (Brogi, 2001; Mazzarone et al., 2000; Perco, 1995). Si può anche aggiungere come la situazione ideale si riscontri in prevalenza su terreni calcarei, calcareo-marinosi ed argillosi, dove la vegetazione caratteristica di tali zone soddisfa le esigenze della specie in Sali minerali. Il capriolo si configura come una specie ubiquitaria e relativamente adattabile ai nuovi ambienti agricoli e boschivi creati dall’uomo, limitatamente a quelle località in cui perdura un agricoltura di tipo tradizionale e abbastanza rispettosa dell’ambiente. Infatti lo possiamo trovare frequentemente in cedui abbandonati e in piccoli boschetti a “macchia di leopardo”, alternati a coltivi e prati. La specie mantiene comunque uno stretto legame con il bosco, anche in aree dove la copertura arborea è scarsa perché, nonostante la quantità di offerta trofica presente nelle aree aperte, queste non rispondono alle esigenze di protezione degli animali, che infatti svolgono le fasi più importanti del loro ciclo biologico proprio in bosco (Hewison et al., 1998; Mazzarone et al., 2000). Il modello di uso dell’habitat del capriolo è condizionato principalmente dalla presenza di ambienti in grado di offrire un nutrimento qualitativamente ricco e ben distribuito durante tutto l’anno, in virtù della sua marcata indole territoriale e stanziale. Le dimensioni dell’ home range (area familiare) dipendono dalla stagione, dalle disponibilità alimentari, dall’età e dal sesso degli animali. Negli ambienti tipici della specie (bosco di latifoglie con ricco sottobosco e ampie radure o aree incolte di pascolo) l’home range varia dai 20 ai 50 ha, sebbene in aree fortemente coltivate senza copertura possa raggiungere i 150-500 ha (Ferloni, 1998; Liberg et al., 1998; Sempéré et al., 1996). Normalmente, a causa dei continui spostamenti alla ricerca di cibo, il domicilio invernale è maggiore rispetto a quello primaverile-estivo (può essere dalle 3 alle 15 volte maggiore rispetto all’area delimitata dal maschio in fase territoriale), e la bassa aggressività degli animali in questo periodo permette una parziale sovrapposizione degli home range (Cederlund, 1983). Il territorio dei maschi dominanti è una zona interna all’home range stagionale, delimitata da confini precisi e poco mutevoli, che risulta relativamente stabile nel tempo. Le dimensioni di quest’area possono variare dai 2 ai 200 ha, ed aumentano al diminuire della densità della popolazione e della qualità dell’ambiente estivo (Liberg et al., 1998; Sempéré et al., 1996). Il territorialismo maschile è molto importante per la dinamica della popolazione, perché favorisce due fenomeni: 1) la colonizzazione dei territori limitrofi ad un nucleo in cui si sia formata una popolazione vitale, a causa delle migrazioni a cui sono costretti i giovani maschi. 2) l’autoregolazione della densità di popolazione in quanto un determinato comprensorio non può contenere più di tanti maschi territoriali. Tra un territorio e l’altro esistono comunque delle zone “cuscinetto”, entro le quali l’aggressività è ridotta al minimo e dove i maschi di territori confinanti riescono a tollerarsi a vicenda (Tarello, 1991). Anche la femmina prossima a partorire assume un comportamento territoriale, in quanto si isola in un luogo tranquillo, di norma non più grande di 2 ha, caratterizzato da microclima favorevole ed in cui può esercitare un controllo sull’ambiente circostante (Hewison et al., 1998; Perco, 1995). Dopo un periodo relativamente breve successivo al parto, l’area vitale aumenta di nuovo e si può sovrapporre a quella di altre femmine (Danilkin, 1996).
Alessandro Cerofolini, laureato in Scienze Faunistiche presso la Facoltà di Agraria di Firenze, frequenta l’ultimo anno del corso di laurea magistrale in scienze faunistico ambientali. Curriculum vitae >>>
 |
Fauna delle Alpi – Mammiferi 87 specie di mammiferi selvatici presenti nelle Alpi italiane, illustrate e dettagliatamente descritte in altrettante schede. 230 fotografie a colori per riconoscere le specie trattate. |





